| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 1g |
|
||
| 2g |
|
||
| 10g |
|
||
| 25g |
|
||
| 50g |
|
||
| 100g |
|
||
| 200g |
|
||
| Other Sizes |
|

| 靶点 |
Microbial Metabolite; Endogenous Metabolite
|
||
|---|---|---|---|
| 体外研究 (In Vitro) |
生长素是一种重要的植物激素,参与多种植物发育过程。生长素水平的时空调节对于在适当的时间和地点实现器官的发育是必要的。这些水平可以通过生长素[吲哚3-乙酸(IAA)]从其共轭形式及其前体的转化来调节。Indole-3-butyric acid/吲哚-3-丁酸(IBA)是一种生长素前体,在过氧化物酶体β氧化过程中转化为IAA。在拟南芥中,IBA转化为IAA的改变会导致多种植物缺陷,表明IBA在关键方面有助于生长素的稳态。与IAA一样,IBA及其结合物可以在植物中运输,但许多IBA载体仍需鉴定。在这篇综述中,我们讨论了迄今为止在拟南芥中鉴定的IBA转运蛋白,包括ATP结合盒转运蛋白(ABCG)家族G亚家族的多效耐药性(PDR)成员,主要促进因子超家族(MFS)家族的IBA1转运蛋白(TOB1)成员,并假设其他潜在的IBA载体参与植物发育。[1]
在培养开始时,两种生长素的水平都无法检测到,这表明拟南芥TCL是研究外源生长素完全控制下AR形成的最佳选择。分析了不同生态型、转基因系和敲除突变体的TCL在不同处理下的AR反应。研究表明,Indole-3-butyric acid/吲哚-3-丁酸/IBA比IAA和IBA+Kin更能诱导AR。IBA诱导的IAA外排(PIN1)和IAA内流(AUX1/LAX3)基因、IAA内流载体活性以及参与IAA生物合成的ANTHRANILATE合成酶-α1(ASA1)基因的表达。ASA1和ANTHRANILATE合成酶β1(ASB1)是同一酶的另一个亚基,在外源IBA存在的情况下,它们对AR的形成有积极影响,因为它们的突变体wei2wei7的TCL中的AR反应大大降低。来自ech2ibr10突变体的经IBA处理的TCL的AR反应也显著降低,该TCL被阻断为IBA到IAA的转化。一氧化氮是IAA下游信号,也是IBA转化为IAA的副产物,在IAA和IBA处理的TCL中早期检测到,但在后一种外植体中检测到的水平更高。 结论:总之,研究结果表明,吲哚-3-丁酸通过转化为涉及NO活性的IAA,以及对IAA转运和ASA1/ASB1介导的IAA生物合成的积极作用,诱导AR的形成。这些结果对于旨在克服具有经济价值的物种的生根阻力的应用非常重要,但主要是为了帮助理解IBA在不定根自然过程中的作用。[2] 受控的植物生长需要通过各种信号分子进行调节,包括类固醇、肽、氧和氮自由基,以及“经典”的植物激素组。生长素对控制植物生长至关重要,也协调许多发育过程,如新根的形成。它通过转录水平的作用缓慢调节根结构,更快速地通过主要靶向质膜感觉系统和细胞内信号通路的机制调节根结构。后一种反应使用几种第二信使,包括Ca(2+)、一氧化氮(NO)和活性氧(ROS)。在这里,我们研究了两种生长素,主要生长素吲哚-3-乙酸(IAA)和另一种内源性生长素Indole-3-butyric acid/吲哚-3-丁酸(IBA)在拟南芥和玉米侧根形成过程中的不同作用。这主要通过不同类型的荧光显微镜和NO产生抑制剂进行分析。这项研究表明,过氧化物酶体IBA转化为IAA后是过氧化物酶体制NO,这对IBA诱导的侧根形成很重要。我们得出结论,过氧化物酶体NO在生长素诱导的根器官发生中起着新的作用。特别是,过氧化物酶体中NO和IAA的空间和时间协调释放是IBA强烈促进侧根形成的原因。[3] 体外活动:在一年中的良好产蛋季节,雏鸡的总产量约为 80% 至 85%,可孵化蛋的百分比高达约 80% 至 90%。在一年中产蛋不佳的季节,雏鸡的总产量约为 50% 至 55%,而可孵化蛋的百分比高达约 60% 至 65%。在运行良好的商业孵化场中,任何一年的平均孵化率约为孵化蛋总数的 75%,这被认为是优秀的。细胞测定:对于自然受精且之前没有活胚胎迹象的卵子,用最佳量的Indole-3-butyric acid/IBA处理促进了某些卵子中活胚胎的形成。它进一步提高了特定批次鸡蛋的雏鸡产量。在卵子胚层的赋予生命的细胞中,最佳量的IBA刺激了细胞的生命生长活性,并导致活胚胎的形成,尽管在最仔细的照光处理之前,该形成未能显现出来。 |
||
| 体内研究 (In Vivo) |
对于受精禽蛋的活体动物胚胎来说,当提供Indole-3-butyric acid/IBA时,它会刺激孵化过程中的生物生命生长过程。在所有情况下,经过 IBA 治疗后,孵出的雏鸡都健康、有活力且充满活力。它们比未经处理的鸡蛋孵出的普通小鸡更能抵抗疾病
|
||
| 细胞实验 |
TCL cultyre[2]
从花序茎的节间切下由包括表皮在内的六层细胞组成的约0.5×8mm的浅表TCL。在22±2°C的持续黑暗中,在由添加了0.55 mM肌醇、0.1μM硫胺素HCL、1%(w/v)蔗糖、0.8%琼脂(w/v,pH 5.7)的MS盐组成的培养基上培养TCL,表皮朝上,直至第15天。Col-0 TCL在该培养基上培养,添加10μM IBA/吲哚-3-丁酸、10μM IBA加0.1μM Kin、0.1μM Kin、10μM IAA、10μM-IBA加0.01μM MeJA,并在HF下作为实验对照。将ech2ibr10、wei2-1wei7-1和lax3aux1-21突变体及其野生型的TCL与10μM IBA、10μM IAA或在HF下培养。ASA1::GUS TCL用10μM IPA或10μM NAA或IBA加0.01μM MeJA培养。用10μM IBA培养DR5::GUS、PIN1::GUS和LAX3::GUS以及Col和Col-gl1生态型的TCL。每个基因型和每个重复使用100个外植体。在高压灭菌之前,用1M NaOH将pH值调节至5.7。对于宏观分析,在培养结束时,在LEICA MZ8立体显微镜下检查WT和突变体的外植体,并将AR反应评估为培养结束时留在初始阶段或形成宏观愈伤组织和AR的外植体百分比,以及每个生根外植体的平均AR数(±SE)。 GUS活性的组织化学分析[2] 在培养的第8天和第15天收获DR5::GUS、PIN1::GUS和LAX3::GUS,AUX1::GUS的TCL,并按照Willemsen等人的描述用GUS染色进行处理,如Veloccia等人所述,进行了细微的修改。在真空钟形罩中渗透15分钟后,将样品在37°C的黑暗中孵育30分钟(DR5:∶GUS和LA X3::GUS),或45分钟(AUX1:∶GUS,ASA1::GUS),或2.5小时(PIN1:∶GUS)。)。GUS测定后,将样品固定在70%(v/v)乙醇中,用分级乙醇系列脱水,包埋在Technovit 7100中,用Microm HM 350 SV切片机纵向切片12μm,并在光学显微镜下观察。 激素定量[2] 在时间0(即切除后不久)收集Col-0的TCL,并保存至-80°C,直至分析。根据Veloccia等人的研究,使用50mg TCL的等分试样提取IAA和IBA/吲哚-3-丁酸。通过快速分辨反相HPLC(RR-RP-HPLC)分离进行IAA和IBA的定量测定,然后用带有ESI界面的三重四极杆(QqQ)质谱仪进行MS/MS检测。两种生长素的纯标准品、内标品和定量依据Veloccia等人。 一氧化氮检测[2] 在落射荧光显微镜下,使用可渗透细胞的二乙酸酯衍生物二氨基荧光素FM(DAF-FMDA)定量用10μM Indole-3-butyric acid/IBA或10μM IAA培养的Col-0 TCL中的细胞内NO含量。在确认仅用缓冲液检测不到明显的落射荧光信号后,在添加了5μM DAF-FMDA的20 mM HEPES/NaOH缓冲液(pH 7.4)中培养2、3和6天,将TCL孵育20分钟(附加文件1:图S1a-b)。用缓冲液洗涤三次以去除多余的荧光探针后,使用配备特定滤光片组(EX 450–490、DM 510、LP 515)的徕卡DMRB显微镜观察TCL。这些图像是用LEICA DC500数码相机采集的,并用IM1000图像分析软件进行分析。每次处理20个TCL,随机进行10次观察,使用ImageJ软件量化荧光信号(绿色)的强度,并以任意单位(AU;0至255)表示。将这些值取平均值并归一化为对照值,即在没有荧光探针的缓冲液中孵育的TCL中测量的值。 将野生型和lrt1突变体的玉米粒(Zea mays L.)浸泡6小时,在室温下在黑暗中在湿润的滤纸卷上发芽4天。选择长度为50-70mm的直主根幼苗(野生型,lrt1)进行生长素处理。在药理学实验中,根尖在室温下在黑暗中浸入适当的溶液中。对于用生长素处理根,在将根尖浸没2小时之前,立即制备10μM IAA或IBA/吲哚-3-丁酸的有效工作浓度。 拟南芥种子经过表面灭菌处理后,放置在不含维生素且含有1%蔗糖(β-氧化缺陷突变体和相关野生型对照为1.5%)的半强度Murashige&Skoog(1962)(MS)培养基上,该培养基用0.8%植酸酶固化。将装有种子的盘子在4°C下储存48小时以打破休眠,然后在连续的黄光下垂直安装3-4天。 拟南芥突变体pex5-1 pex7-1、pxa1-1和pex6(最初由Zolman等人于2000年指定为B11)具有β-氧化缺陷,并显示出IBA/吲哚-3-丁酸不敏感性(Zolman等,2001a;Zolman&Bartel,2004;Woodward&Bartel),以及NO产生受损的突变体noa1(最初由Guo等人于2003年指定为Atnos1)用于我们的实验。 为了进行显微镜观察,将3-4d龄的幼苗转移到显微镜载玻片上,将其放置在由盖玻片制成的薄室中。将腔室充满半强度MS培养基,但不含植脂凝胶,并将其放置在无菌玻璃试管中,试管中的培养基水平达到腔室的开口下端。这使得腔室和反应杯之间可以自由交换介质。幼苗在连续的黄光下垂直生长长达24小时。在此期间,幼苗在液体培养基中稳定了根系生长并产生了新的根毛。使用过氧化物酶体靶向信号1-绿色荧光蛋白(PTS1-GFP)对过氧化物酶体内进行监测(参见Woodward&Bartel,2005b)。 |
||
| 动物实验 |
|
||
| 毒性/毒理 (Toxicokinetics/TK) |
非人类毒性值
小鼠口服LD50 100 mg/kg 小鼠腹腔注射LD50 100 mg/kg 8617 大鼠口服LD >500 mg/kg 美国国家科学院、国家研究委员会、化学-生物协调中心,《评论》,5(7),1953 8617 小鼠口服LD50 100 mg/kg 《农业化学品》,Thomson, WT,4卷,加利福尼亚州弗雷斯诺,Thomson出版社,1976/77修订版,3(76),1976/1977 8617 小鼠腹腔注射LD50 100 mg/kg 《农业化学品手册》(含更新版),Hartley, D.和H. Kidd主编,诺丁汉,皇家化学学会, 1983-86,A231(1983) |
||
| 参考文献 |
[1]. Indole 3-Butyric Acid Metabolism and Transport in Arabidopsis thaliana. Front Plant Sci. 2019 Jul 3;10:851.
[2]. Indole-3-butyric acid promotes adventitious rooting in Arabidopsis thaliana thin cell layers by conversion into indole-3-acetic acid and stimulation of anthranilate synthase activity. BMC Plant Biol. 2017 Jul 11;17(1):121. [3]. Indole-3-butyric acid induces lateral root formation via peroxisome-derived indole-3-acetic acid and nitric oxide. New Phytol. 2013 Oct;200(2):473-482. |
||
| 其他信息 |
吲哚-3-丁酸(IBA)是一种吲哚-3-基羧酸,它是丁酸在1位带有1H-吲哚-3-基取代基的化合物。它是一种植物激素、植物代谢物和生长素。其功能与丁酸相关,是吲哚-3-丁酸酯的共轭酸。
据报道,玉米(Zea mays)、椰子(Cocos nucifera)和其他一些有相关数据的生物体中都存在吲哚-3-丁酸。 IBA是拟南芥三叶草(TCLs)不定根形成的主要促进因子,也可能在许多其他内源生长素含量极低的培养体系和物种中发挥重要作用。IBA通过转化为IAA,并增强IAA的生物合成和运输发挥作用。其作用的关键在于调节内源IAA库。 IBA对IAA稳态的调控涉及其过氧化物酶体转化下游的其他化合物,如NO和茉莉酸酯的活性。IBA与NO和茉莉酸酯的关系,以及下游生长素信号传导和感知机制,仍需进一步研究。尽管这些发现有助于设计实验以克服具有经济价值的物种的生根困难,但其主要意义在于帮助理解IBA控制不定根自然形成过程的机制[2]。内源IBA的作用仍有待解答。近期研究表明,IBA向IAA的转化通过补充内部活性生长素库,对幼苗的正常发育至关重要(Strader等,2011)。许多无法利用IBA的突变体的一个表型特征是侧根数量不足(例如Zolman等人,2001a,b;Wiszniewski等人,2008;Strader等人,2010, 2011)。这可能表明IBA转化为IAA对于侧根生长(LRF)是必需的。我们目前的数据表明,NO和IAA在空间和时间上的协调释放是IBA生物活性所必需的。IBA在过氧化物酶体中转化为IAA不仅导致IAA合成受到严格调控,而且还促进NO的生成。可以预期,其他同样需要IBA且涉及NO的过程也将依赖于过氧化物酶体中IAA-NO的生成。在此,我们可以提及根毛的极化顶端生长以及根对盐胁迫和水分胁迫的适应性(Lombardo et al., 2006; Strader et al., 2010, 2011; Tognetti et al., 2010; Strader & Bartel, 2011)。未来的研究需要阐明哪些类型的NO生成酶系统负责IBA诱导的NO生成,以及NO介导的信号传导是否也参与IBA发挥作用或必不可少的其他过程。[3] |
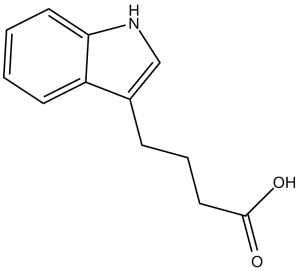
| 分子式 |
C12H13NO2
|
|
|---|---|---|
| 分子量 |
203.24
|
|
| 精确质量 |
203.094
|
|
| 元素分析 |
C, 70.92; H, 6.45; N, 6.89; O, 15.74
|
|
| CAS号 |
133-32-4
|
|
| 相关CAS号 |
|
|
| PubChem CID |
8617
|
|
| 外观&性状 |
White to slightly yellow crystals
White to tan powder or crystalline solid |
|
| 密度 |
1.3±0.1 g/cm3
|
|
| 沸点 |
426.6±20.0 °C at 760 mmHg
|
|
| 熔点 |
124-125.5 °C(lit.)
|
|
| 闪点 |
211.8±21.8 °C
|
|
| 蒸汽压 |
0.0±1.1 mmHg at 25°C
|
|
| 折射率 |
1.646
|
|
| LogP |
2.34
|
|
| tPSA |
53.09
|
|
| 氢键供体(HBD)数目 |
2
|
|
| 氢键受体(HBA)数目 |
2
|
|
| 可旋转键数目(RBC) |
4
|
|
| 重原子数目 |
15
|
|
| 分子复杂度/Complexity |
230
|
|
| 定义原子立体中心数目 |
0
|
|
| SMILES |
O([H])C(C([H])([H])C([H])([H])C([H])([H])C1=C([H])N([H])C2=C([H])C([H])=C([H])C([H])=C12)=O
|
|
| InChi Key |
JTEDVYBZBROSJT-UHFFFAOYSA-N
|
|
| InChi Code |
InChI=1S/C12H13NO2/c14-12(15)7-3-4-9-8-13-11-6-2-1-5-10(9)11/h1-2,5-6,8,13H,3-4,7H2,(H,14,15)
|
|
| 化学名 |
4-(1H-indol-3-yl)butanoic acid
|
|
| 别名 |
|
|
| HS Tariff Code |
2934.99.9001
|
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
|
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
|
|||
|---|---|---|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.5 mg/mL (12.30 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 25.0 mg/mL 澄清 DMSO 储备液加入900 μL 玉米油中,混合均匀。 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 4.9203 mL | 24.6015 mL | 49.2029 mL | |
| 5 mM | 0.9841 mL | 4.9203 mL | 9.8406 mL | |
| 10 mM | 0.4920 mL | 2.4601 mL | 4.9203 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 COA
COA
 463611831
463611831
