| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 100mg |
|
||
| 250mg |
|
||
| 500mg |
|
||
| 5g |
|
||
| Other Sizes |
|

| 靶点 |
Apical membrane antigen 1 (AMA1) of Plasmodium falciparum. For the N-Me-Leu-8 peptide (synthesized using Fmoc-N-Me-Leu-OH):
- Binding affinity (KD) to 3D7 strain AMA1: 21.00 ± 1.73 nM (kinetic analysis) [1] - Binding affinity (KD) to W2mef strain AMA1: 16.8 ± 0.1 μM (steady-state analysis) [1] - Binding affinity (KD) to HB3 strain AMA1: 68.5 ± 0.5 μM (steady-state analysis) [1] |
|---|---|
| 体外研究 (In Vitro) |
抑制寄生虫入侵:N-Me-Leu-8肽(含N-甲基亮氨酸)对3D7型恶性疟原虫入侵红细胞的抑制作用强于天然R1肽。在50 μg/mL浓度下,N-Me-Leu-8肽可完全抑制寄生虫入侵。滴定实验表明,N-Me-Leu-8肽的入侵抑制作用强于天然R1肽,这与其增强的AMA1亲和力相关。[1]
- AMA1结合(ELISA):N-Me-Leu-8肽与R1噬菌体竞争结合固定化的3D7 AMA1。其IC50值(抑制R1噬菌体结合50%的浓度)低于天然R1肽,表明其结合能力更强。 [1] - 在小鼠血浆中的蛋白水解稳定性:N-Me-Leu-8肽与天然R1相比,稳定性显著提高。在37°C的小鼠血浆中孵育80分钟后,约50%的N-Me-Leu-8肽仍保持完整,而天然R1在10分钟内几乎完全降解。[1] |
| 酶活实验 |
表面等离子共振(SPR)结合分析:使用Biacore T100生物传感器在25°C下测定了N-Me-Leu-8肽与不同恶性疟原虫株(3D7、HB3、W2mef)AMA1蛋白的相互作用动力学。AMA1通过标准胺偶联法固定在CM5传感器芯片上。运行缓冲液为HBS-EP+(10 mM HEPES、150 mM NaCl、3.4 mM EDTA、0.05%表面活性剂P20,pH 7.4)。将浓度范围为 5 至 320 nM(针对 3D7)或 0.32 至 40.96 μM(针对 W2mef 和 HB3)的肽溶液以 60 μL/min 的流速注入固定化的 AMA1 上 1 分钟,随后进行 5 分钟的解离。使用 Scrubber 软件处理传感图。通过对包含质量传递项的 1:1 结合模型进行全局拟合来确定速率常数(ka 和 kd)。KD 计算为 kd/ka。对于快速解离的相互作用,采用稳态亲和力分析。对于N-Me-Leu-8与3D7 AMA1的结合:ka = (2.74 ± 0.54)×10^7 1/Ms,kd = (2.84 ± 0.15)×10^-2 1/s,KD = 21.00 ± 1.73 nM。[1]
- 基于ELISA的竞争性结合实验:将96孔板用2 μg/mL重组AMA1(3D7菌株)于4°C包被过夜。用10%脱脂奶粉PBS溶液封闭后,向孔中加入不同浓度的N-Me-Leu-8肽以及恒定浓度的R1展示噬菌体(~2×10^9 cfu/mL)。室温下振荡孵育 1 小时后,用 PBS/0.05% Tween 20 洗涤去除未结合的噬菌体。使用辣根过氧化物酶标记的抗 M13 抗体(1:5000)和四甲基联苯胺底物检测结合的噬菌体,并在 450 nm 处读取吸光度。测定使 R1-噬菌体结合减少 50% 所需的肽浓度(IC50)。[1] |
| 细胞实验 |
入侵抑制试验(寄生虫生长抑制):将3D7寄生虫株培养并用山梨醇裂解法同步化至裂殖体期。将肽(包括N-Me-Leu-8)用PBS稀释,取50 μL加入无菌平底微孔板中,每个样品重复三次。然后加入50 μL完全培养基和100 μL同步化的裂殖体期寄生虫(4%红细胞比容,0.3%寄生虫血症)。将微孔板置于37℃、94% N₂、1% O₂、5% CO₂的湿润环境中培养40-42小时。用冰冷的PBS洗涤后,将寄生虫冻融,并通过测定寄生虫乳酸脱氢酶活性来确定相对寄生虫血症。在650 nm处读取吸光度值。抑制率计算公式为:100 − [(A650 肽样品 − A650 仅 RBC) / (A650 无肽对照 − A650 仅 RBC) × 100]。在滴定实验中,N-Me-Leu-8 的效力较天然 R1 有所提高(浓度-反应曲线见图 3C)。[1]
|
| 药代性质 (ADME/PK) |
小鼠血浆中的蛋白水解稳定性:将N-Me-Leu-8肽(0.4 mg)溶解于含0.1%苯甲醇(作为内标)的PBS缓冲液(400 μL)中。将等体积的小鼠血浆溶液(由冻干血浆复溶)和肽溶液混合(总体积800 μL),并在37°C下孵育。分别于0、10、20、40和80分钟取样(50 μL),每个时间点重复三次,置于冰上,加入20 μL 0.5 M赖氨酸盐酸盐和65 μL乙腈,冷却后离心,取上清液进行反相高效液相色谱分析。肽峰面积与苯甲醇峰面积的比值以t=0时点为基准进行归一化。 N-Me-Leu-8表现出显著提高的稳定性:80分钟后,约50%的肽仍保持完整,而天然R1在10分钟内几乎完全降解(剩余<20%)。[1]
|
| 参考文献 | |
| 其他信息 |
背景:R1肽是一种由20个氨基酸残基组成的疟原虫入侵抑制剂,可与AMA1结合。然而,它易被蛋白酶快速降解,且具有菌株特异性活性。研究人员探索了N-甲基化修饰,包括在Leu-8位使用Fmoc-N-Me-Leu-OH进行修饰,以提高其稳定性、亲和力和特异性广度。[1]
- 作用机制:N-Me-Leu-8肽保留了与AMA1结合并阻断寄生虫入侵的能力。亲和力的提高(解离速率减慢)表明N-甲基化修饰稳定了肽-AMA1复合物。该肽可能靶向AMA1上一个保守的疏水口袋,因为它能与抑制性单克隆抗体1F9和4G2dc1竞争结合位点。[1] - 结构影响:核磁共振研究表明,Leu-8位的N-甲基化修饰不会引起长程结构变化;骨架化学位移差异是局部的。N-Me-Leu-8的溶液结构与天然R1相似,具有两个转角区域。然而,N-甲基NOE的引入表明其具有一定的局部稳定性。[1] - 菌株特异性:天然R1仅与3D7和D10菌株结合,而N-Me-Leu-8与HB3和W2mef AMA1表现出微弱但可测量的结合(KD分别为~68.5 μM和~16.8 μM),表明其对不同菌株的识别能力有所提高。 |
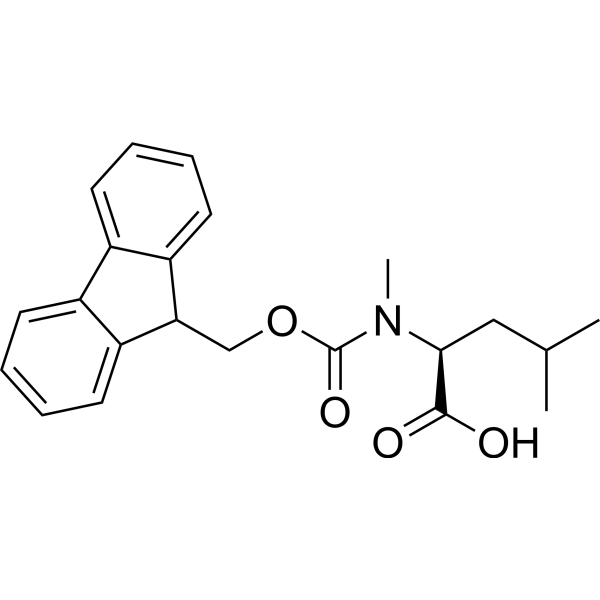
| 分子式 |
C22H25NO4
|
|---|---|
| 分子量 |
367.4382
|
| 精确质量 |
367.178
|
| CAS号 |
103478-62-2
|
| PubChem CID |
7015835
|
| 外观&性状 |
White to off-white solid powder
|
| 密度 |
1.2±0.1 g/cm3
|
| 沸点 |
537.3±29.0 °C at 760 mmHg
|
| 熔点 |
113-116ºC
|
| 闪点 |
278.7±24.3 °C
|
| 蒸汽压 |
0.0±1.5 mmHg at 25°C
|
| 折射率 |
1.581
|
| LogP |
5.44
|
| tPSA |
66.84
|
| 氢键供体(HBD)数目 |
1
|
| 氢键受体(HBA)数目 |
4
|
| 可旋转键数目(RBC) |
7
|
| 重原子数目 |
27
|
| 分子复杂度/Complexity |
512
|
| 定义原子立体中心数目 |
1
|
| SMILES |
CC(C)C[C@@H](C(=O)O)N(C)C(=O)OCC1C2=CC=CC=C2C3=CC=CC=C13
|
| InChi Key |
BUJQSIPFDWLNDC-FQEVSTJZSA-N
|
| InChi Code |
InChI=1S/C22H25NO4/c1-14(2)12-20(21(24)25)23(3)22(26)27-13-19-17-10-6-4-8-15(17)16-9-5-7-11-18(16)19/h4-11,14,19-20H,12-13H2,1-3H3,(H,24,25)/t20-/m0/s1
|
| 化学名 |
(2S)-2-[9H-fluoren-9-ylmethoxycarbonyl(methyl)amino]-4-methylpentanoic acid
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO : ~100 mg/mL (~272.15 mM)
|
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.5 mg/mL (6.80 mM) (饱和度未知) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 25.0 mg/mL澄清DMSO储备液加入到400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: 2.5 mg/mL (6.80 mM) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (这些助溶剂从左到右依次添加,逐一添加), 悬浊液; 超声助溶。 例如,若需制备1 mL的工作液,可将 100 μL 25.0 mg/mL澄清DMSO储备液加入900 μL 20% SBE-β-CD生理盐水溶液中,混匀。 *20% SBE-β-CD 生理盐水溶液的制备(4°C,1 周):将 2 g SBE-β-CD 溶解于 10 mL 生理盐水中,得到澄清溶液。 View More
配方 3 中的溶解度: ≥ 2.5 mg/mL (6.80 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.7215 mL | 13.6077 mL | 27.2153 mL | |
| 5 mM | 0.5443 mL | 2.7215 mL | 5.4431 mL | |
| 10 mM | 0.2722 mL | 1.3608 mL | 2.7215 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 463611831
463611831
