| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| 500mg |
|
||
| Other Sizes |
|
| 靶点 |
ROS; mitochondria-targeted antioxidant
|
|---|---|
| 体外研究 (In Vitro) |
线粒体醌 (MitoQ) 是一种针对线粒体的抗氧化剂。在四个小时的冷藏 (CS) 期间,使用剂量反应测试来确定米托醌 (MitoQ) 和 DecylTPP 治疗的理想剂量。为了研究线粒体醌治疗对 CS 损伤的潜在保护作用,首先使用了 MitoSOX Red(一种靶向并检测线粒体超氧化物产生的荧光染料)。当CS应用于正常大鼠肾(NRK)细胞时,线粒体超氧化物导致细胞荧光比对照组增加约两倍。当涉及 CS 诱导的线粒体超氧化物产生时,线粒体中的醌可以显着抵抗它,而对照化学物质 DecylTPP 则没有这种作用。线粒体超氧化物治疗大大减少了线粒体超氧化物的产生,并且用 DecylTPP 治疗的肾脏的线粒体超氧化物水平与仅接受 CS 的肾脏相当[1]。
MitoQ对CS期间线粒体超氧化物生成的保护作用。[1] MitoQ和DecylTPP治疗的最佳剂量是从4-h CS期间的剂量反应实验中选择的。对于基于细胞的研究,从0.5、0.75、1.0、1.5和2μM的范围内选择1μM,而离体研究的最有效剂量是50、100和500μM的剂量反应研究中的100μM(数据未显示)。最初使用线粒体靶向荧光染料MitoSOX Red测试了MitoQ治疗对CS损伤的潜在保护作用,该染料可测量线粒体超氧化物的产生。如图2A所示,与未处理的细胞相比,暴露于CS的NRK细胞由于线粒体超氧化物而导致荧光增加了约2倍。MitoQ对CS诱导的线粒体超氧化物产生具有显著的保护作用;而对照化合物DecylTPP没有提供任何保护。使用基于荧光分光光度法的测定法在396和510 nm处检测MitoSOX Red荧光激发,进一步证实了这一点,其中396 nm是线粒体超氧化物的特异性指示剂,510 nm检测非特异性氧化剂的产生(Robinson等人,2006)(图2B)。此外,与对照组肾脏相比,单独暴露于CS的肾脏显示线粒体超氧化物生成增加(图2C)。MitoQ治疗显著降低了线粒体超氧化物的产生,而用DecylTPP治疗的肾脏的线粒体超氧化物水平与单独暴露于CS的肾脏相当(图2C)。 MitoQ减弱CS过程中硝基酪氨酸加合物的形成。[1] 免疫细胞化学和免疫组织化学用于评估CS期间的硝基酪氨酸蛋白加合物。如图3A所示,与未处理的细胞相比,单独暴露于CS的NRK细胞的硝基酪氨酸染色(红色荧光)显著增加。MitoQ将硝基酪氨酸的形成减少了2倍,而对照化合物DecylTPP没有减少CS介导的硝基酪氨酸的生成(图3A)。过氧化亚硝酸盐处理的细胞具有强烈的硝基酪氨酸形成(阳性对照),当硝基酪氨酸抗体被过量3-硝基酪氨酸预吸收时被阻断(阴性对照;ONOO-+阻断)。大鼠肾脏免疫组织化学数据与关于MitoQ和硝基酪氨酸作用的体外研究结果一致。图3B显示,与对照组肾脏相比,CS肾脏的远端和近端小管中硝基酪氨酸(棕色染色)的增加,肾小球中的增加程度较小。用MitoQ治疗的肾脏硝基酪氨酸形成较少。相比之下,DecylTPP治疗的肾脏与CS肾脏的硝基酪氨酸含量相似。使用用过量3-硝基酪氨酸预吸收的抗体(CS+阻断)也证实了硝基酪氨酸染色的特异性。 MitoQ防止CS期间线粒体呼吸复合物失活。[1] 在体外和离体肾模型中评估线粒体呼吸复合物活性,以研究4-h CS是否会改变线粒体呼吸功能。与未处理的细胞相比,NRK细胞CS后复合物I和II显著失活(图4)。MitoQ完全阻止了复合物I和II的失活,而DecylTPP对复合物活性没有显著影响。本研究未对复合物III和IV进行评估,因为我们之前已经证明,24小时CS对这两种复合物的活性都没有影响(Mitchell等人,2010);因此,没有必要对这些复杂的活动进行评估。与体外研究结果一致,大鼠肾脏的CS导致复合物I和II活性的部分失活,对复合物III和IV没有影响(图4)。MitoQ对CS肾脏的复合物I和II失活具有保护作用,而DecylTPP没有任何作用。 MitoQ减少NRK细胞CS+再武装过程中硝基酪氨酸的形成和细胞死亡。[1] 为了测试MitoQ是否可能在再灌注/移植后提供对氧化剂产生和细胞死亡的保护,细胞暴露于CS加RW。如图6A所示,NRK细胞的4-h CS加过夜(18小时)RW导致显著的硝基酪氨酸形成。在CS期间添加MitoQ显著减弱了CS期间的硝基酪氨酸染色。而对照化合物DecylTPP没有效果。LDH细胞毒性显示,暴露于CS的NRK细胞的细胞死亡显著增加。RW与未处理细胞的比较(图6B)。MitoQ在单独CS和CS期间将细胞死亡降低了约2倍。RW.DecylTPP在两个治疗组中都没有减少细胞死亡。 MitoQ清除分离的胰腺腺泡细胞中的ROS产生[2] 应用1 mM H2O2导致细胞(对照组)中ROS的稳定上升,这反映在CM-H2DCFDA荧光强度的增加上;用1预处理的细胞 μM MitoQ与对照组细胞相比,ROS产生显著减少,而1 μM dTPP,没有抗氧化活性的MitoQ亲脂性阳离子没有影响(图1(a))。MitoQ和dTPP本身均未诱导ROS的产生(图1(b))。 MitoQ不能保护分离的胰腺腺泡细胞免受AP沉淀剂引起的线粒体去极化[2] 未用1进行预处理 μM MitoQ和dTPP在分离的胰腺腺泡细胞中引起了ΔΨM的去极化,这与实验结束时应用原核CCCP以引发完全去极化形成鲜明对比。(图2(a))。然而,在10 μM的MitoQ和dTPP都诱导了ΔΨM本身的稳定降低,这对MitoQ来说更为深刻(图2(b)和2(c))。与单独用HEPES处理的对照细胞相比,CCK(10 nM)诱导了ΔΨm的去极化(图2(d)),这种作用不受1 μM MitoQ或dTPP(图2(e))。同样,TLCS(500μM)的灌注使ΔΨM去极化(图2(f)),而1不受影响 μM MitoQ或dTPP(图2(g))。 MitoQ导致胰腺腺泡细胞死亡并加重CCK诱导的坏死[2] 两者1 μM MitoQ和dTPP引起PI摄取增加,表明坏死;在2、6和10时观察到显著差异 h,用MitoQ或dTPP预处理的细胞与单独用HEPES处理的对照组细胞之间的差异(图3(a))。增加10 nM CCK诱导坏死的时间依赖性增加。然而,1 μM MitoQ对CCK诱导的细胞死亡没有任何保护作用。相反,与单独使用CCK相比,MitoQ和dTPP在2℃时均显著恶化了坏死 h、 而dTPP,而不是MitoQ,在以后的时间点加重了CCK诱导的细胞死亡(图3(b))。 |
| 体内研究 (In Vivo) |
米托醌 (MitoQ) 治疗可显着减少中性粒细胞浸润和胰腺水肿。 MitoQ 会剂量依赖性地增加血清淀粉酶;如果剂量较高,则大约翻倍。当以 10 mg/kg(剂量 1)给药时,MitoQ 治疗显着提高了血清 IL-6 水平,并使 Caerulein 诱导的肺 MPO 活性几乎增加了一倍 [2]。
MitoQ改善了CER-AP的整体胰腺组织病理学,但加重了全身损伤[2] 图4(a)显示了对照组和不同治疗组的代表性组织病理学幻灯片,图4(b)总结了单个成分的总体组织病理学评分和分解评分。腹腔注射生理盐水不会引起胰腺的任何明显的组织病理学变化,而蓝蛙素的过度刺激会引起AP的典型特征;胰腺导管边缘和实质明显水肿、空泡化、中性粒细胞浸润,局灶性腺泡细胞坏死明显12 h第一次注射蛙皮素后。与生理盐水对照组相比,CER-AP的特征还在于血清淀粉酶、胰蛋白酶和MPO活性以及肺MPO活性显著增加。 MitoQ两种剂量的治疗均显著减少了胰腺水肿和中性粒细胞浸润。然而,胰腺坏死没有得到预防,在较高剂量下有更大的坏死趋势,尽管这没有达到显著性。MitoQ剂量依赖性地增加血清淀粉酶,在较高剂量下约加倍(图5(a))。胰蛋白酶活性和MPO活性在任何剂量下都没有受到MitoQ的显著影响(图5(b)和5(c))。此外,MitoQ治疗使蓝蛙素诱导的肺MPO活性几乎翻了一番(图5(d)),在剂量1时,血清IL-6水平也明显升高(图5)。 非抗氧化剂类似物dTPP在两种剂量下均显著减少了水肿、中性粒细胞浸润和坏死,导致组织病理学评分总体降低。尽管dTPP降低了胰蛋白酶和MPO活性(图5(b)和5(c)),但血清淀粉酶没有受到显著影响(图5)。与MitoQ获得的结果类似,dTPP也显著增加了蓝蛙素诱导的肺MPO活性和血清IL-6水平(图5(d)和5(e))。 MitoQ无法抵御TLCS-AP[2] 假手术只引起胰腺腺泡细胞轻度水肿,没有明显的炎症和坏死迹象。输注3 通过胰管进入胰腺的mM TLCS在24小时时导致胰腺头部明显的组织病理学变化 h、 其特征是水肿、炎症、坏死显著增加,从而导致整体组织病理学评分(图6(a)(i-iv))。然而,胰腺的身体和尾部受到的影响要小得多(数据未显示)。与假手术组相比,TLCS-AP与血清淀粉酶、胰腺MPO活性和血清IL-6水平升高有关(图6(b)-6(d))。 较低剂量的MitoQ和dTPP均未诱导胰腺的组织病理学变化,水肿、炎症、坏死和总体组织病理学评分没有变化(图6(a)(i-iv))。同样,当TLCS-AP小鼠用MitoQ或dTPP治疗时,血清淀粉酶和胰腺MPO活性没有显著变化(图6(b)和6(c))。MitoQ或dTPP治疗使血清IL-6水平略有升高,但未达到统计学意义(图6(d))。在没有注射蓝蛙素或输注TLCS的情况下,单独对小鼠应用两种剂量的MitoQ或dTPP表明,MitoQ和dTPP本身都显著增加了肺MPO活性(数据未显示)。 MitoQ对PMA诱导的PMNs中ROS产生的双相效应[2] 图7(a)说明了1的效果 μM MitoQ或dTPP对PMA诱导的分离PMNs中ROS产生的影响。NAD(P)H氧化酶刺激剂PMA(50 ng/mL)在最初的几分钟内诱导PMNs周围细胞外溶液中ROS的急剧增加,在8 大约20分钟后,下降到平稳状态 mins.NAD(P)H氧化酶抑制剂DPI的应用降低了峰值相并完全抑制了ROS平台(图7(c))。MitoQ治疗对PMNs中ROS的产生产生了双相效应。因此,观察到PMA诱导的初始ROS峰值的浓度依赖性抑制,峰值时间延迟至10分钟(图7(a)-7(c))。应用1 与单独用PMA处理的细胞相比,μM dTPP对PMA诱导的PMNs中ROS产生的峰值没有显著影响。有趣的是,MitoQ在40℃时引起PMA诱导的ROS产生的浓度依赖性增强 mins(图7(c)),这是dTPP仅在较高浓度下共享的动作。 MitoQ治疗可减少CS诱导的肾损伤和细胞死亡。[1] 采用过碘酸希夫染色法检测大鼠肾脏CS期间的组织病理学变化。CS暴露后,出现了严重的广泛管状损伤,如扩张、刷状边界损失和细胞碎片/铸型形成(图5A)。MitoQ显著改善了肾脏组织学,而DecylTPP治疗没有逆转肾脏损伤。TUNEL染色,即细胞核的棕色染色,被用作细胞死亡的标志。如图5B所示,与对照组肾脏相比,CS导致细胞死亡(红色箭头)显著增加。与CS肾脏相比,MitoQ治疗将细胞死亡降低了约2倍,而DecylTPP对细胞死亡没有保护作用。 氧化应激引起的线粒体功能障碍可能在睾丸损伤和退化的发展中起关键作用,导致成年后生育能力受损。MitoQ作为线粒体靶向抗氧化剂已在许多疾病中使用了很长时间,但其对睾丸损伤的治疗作用“尚未有报道”。在这里,我们研究了MitoQ对雷公藤甲素(TP)诱导的氧化应激睾丸损伤的保护作用机制。在TP诱导的睾丸损伤模型中,小鼠口服MitoQ(分别为1.3、2.6和5.2mg/kg)14天。然后从形态学变化、精子发生评估、血睾屏障(BTB)完整性和细胞凋亡等方面对睾丸损伤进行综合评估。结果表明,MitoQ有效地增加了睾丸重量,保持了BTB的完整性,通过抑制氧化应激保护了睾丸组织的微观结构和精子形态。进一步的机制研究表明,MitoQ显著激活Keap1-Nrf2抗氧化防御系统,其特征是增加Nrf2及其靶基因HO-1和NQO1的表达。同时,MitoQ上调线粒体动力学蛋白Mfn2和Drp-1的表达,对线粒体具有保护作用。在此基础上,药代动力学研究结果表明,尽管绝对生物利用度较低,但口服后MitoQ可以进入睾丸组织,这为MitoQ治疗睾丸损伤提供了物质基础。更重要的是,MitoQ能够快速到达线粒体,并具有在塞尔托利细胞中靶向线粒体的突出特征。因此,这些结果为MitoQ在睾丸损伤疾病中的应用提供了信息[3]。 |
| 细胞实验 |
体外冷藏模型。[1]
将正常大鼠肾近端肾小管细胞保存在6孔100或150 mm或150 mm板中,置于37°C、含5%胎牛血清(FCS)的DMEM中,用5%CO2和95%空气充气的加湿培养箱中。细胞生长至60%融合,分为四个处理组:1)未处理(Untx),2)CS,3)CS+MitoQ,和4)CS+DecylTPP。未经处理的细胞在含有5%FCS的DMEM中保持在37°C(第1组)。CS是通过用冷PBS洗涤细胞两次并将其单独储存在UW/Viaspan溶液中(4°C下4小时)(第2组)、CS+MitoQ(1μM)(第3组)或CS+DecylTPP(1μM)(第4组)来启动的。在单独的实验中,通过将单独的UW溶液或含有MitoQ或DecylTPP的UW液替换为含有5%FCS的DMEM,将细胞暴露于CS加RW中过夜(37°C下18小时)。 ROS产生的测量[2] 如前所述,使用蔡司LSM510共聚焦显微镜,用探针5-氯甲基-2,7-二氯二氢荧光素二乙酰酯(CM-H2DCFDA)测量胰腺腺泡细胞中的实时ROS产生和氧化还原变化。将新鲜分离的小鼠胰腺腺泡细胞与1 μMMitoQ或dTPP,同时负载10 μM CM-H2DCFDA处理30分钟。然后用H2O2灌注细胞以诱导ROS。CM-H2DCFDA的荧光在488 nm,在505-550处收集发射 nm. 对于中性粒细胞中的ROS测量,使用POLARstar Omega平板读数器进行过氧化物酶增强鲁米诺化学发光测定。细胞以每孔500000的密度铺板,并用1 μMMitoQ或dTPP 10分钟,然后加入50 μM鲁米诺和75单位/mL辣根过氧化物酶。50℃诱导NAD(P)H氧化酶活化 ng/mL肉豆蔻酸佛波醇乙酸酯(PMA),同时使用1 μM二苯碘鎓(DPI)。440处的发光 ROS染料的nm记录为40 min.每只小鼠/每次运行的化学发光强度均归一化为阴性对照。 胰腺腺泡细胞中ΔΨm的测量[2] 在单独的实验中,如前所述,通过四甲基罗丹明甲酯测定法测定胰腺腺泡细胞的ΔΨm。简而言之,这些细胞装载了40 nM TMRM孵育30分钟,然后与1 μM或10 μM的MitoQ或dTPP。胆囊收缩素-8(CCK-8,10 nM)或胆汁酸牛磺胆酸3-硫酸盐(TLCS,500 μM)诱导ΔΨM去极化。灌注结束时,原卟啉羰基氰3-氯苯腙(CCCP,10 μM)诱导ΔΨM完全去极化。TMRM的荧光在543激发 nm,在560-650处收集发射 nm. 胰腺腺泡细胞体外细胞死亡试验[2] 胰腺腺泡细胞死亡是通过坏死细胞核吸收的荧光染料碘化丙啶(PI)的强度来检测的。对于CCK-8诱导的细胞死亡,使用了时间过程荧光板读数法。简而言之,从一只小鼠胰腺中分离细胞,离心,并重新悬浮于1 mL溶液。小心地将细胞移入单个孔中,以确保均匀性。细胞单独用CCK-8(10nM)处理,或在1 μM的MitoQ或dTPP。对于正常对照组,用等体积的细胞外溶液处理细胞。5之后 然后将PI(50μM)加入到通过自动搅拌混合的所有孔中。然后将微孔板放入POLARstar Omega板读数器(预热至37°C)中,通过激发543测定荧光 nm和发射620 nm,底部读数。该测定设置为以600秒的循环时间运行。所有荧光测量值均表示为与基础荧光(F/F 0比)的变化,其中F 0表示实验开始时记录的初始荧光,F表示特定时间点记录的荧光。 |
| 动物实验 |
冷藏离体模型。[1]
体重在250至300克之间的雄性Fischer 344近交系大鼠用乙醚麻醉,随后剃毛并用碘伏消毒。静脉注射2毫升0.9%(w/v)氯化钠溶液,并在耻骨联合上方1厘米至剑突尖端处做切口。用血管钳夹住主动脉和下腔静脉(肾血管近端和远端),以阻断肾脏血流。用22号手术针穿刺大鼠主动脉,通过细导管用生理盐水(每肾10毫升)冲洗肾移植片。一旦肾脏开始变成浅棕色(灌注),在下腔静脉上再开一个孔,允许血液流出肾脏。双肾完全冲洗后,取出右肾作为对照组(第1组),左肾则单独暴露于CS中,于4℃下孵育4小时(第2组)。在后续实验中,使用细导管经主动脉向肾脏灌注生理盐水,随后分别用含MitoQ(100 μM)的生理盐水和含DecylTPP(100 μM)的生理盐水冲洗右肾,用含DecylTPP(100 μM)的生理盐水冲洗左肾(每肾10 ml)。取出组织,分别置于CS + MitoQ(100 μM;右肾)或CS + DecylTPP(100 μM;左肾)中,于4℃下孵育4小时(分别为第3组和第4组)。切取所有肾脏的薄层中部组织,立即用10%福尔马林固定,然后包埋于石蜡中,进行切片(4 μm)和组织学评估。剩余的肾脏组织迅速置于液氮中冷冻,并储存于−80°C直至进行生化分析。 实验性急性胰腺炎模型[2] 每小时腹腔注射一次超最大剂量(50 μg/kg)的CCK-8类似物——胰泌素,共注射7次,以诱导高刺激性急性胰腺炎(CER-AP)。对照组小鼠注射等体积的PBS。在MitoQ治疗组中,分别在第一次和第三次注射胰泌素时给予10 mg/kg(剂量1)或25 mg/kg(剂量2)的MitoQ。同样,在dTPP治疗组中,分别给予9.6 mg/kg(剂量1)或24 mg/kg(剂量2)的dTPP。在剂量 1 和 2 中,MitoQ 和 dTPP 的摩尔浓度相同。小鼠在首次注射胰泌素后 12 小时处死以收集样本。 胆汁酸诱导的急性胰腺炎 (AP) 是通过逆行将 TLCS 注入胰管 (TLCS-AP) 实现的。麻醉诱导后,使用微型输液泵以 5 μL/min 的速度持续输注 TLCS 10 分钟。胰头出现弥漫的浅蓝色(亚甲蓝)表明 TLCS 已成功注入胰腺。对照组小鼠接受假手术,不进行 TLCS 输注。在治疗组中,分别在 TLCS 输注后 1 小时和 3 小时给予 MitoQ (10 mg/kg) 或 dTPP (9.6 mg/kg)。小鼠在 TLCS 输注或假手术后 24 小时处死。在两种动物模型中,均通过给予0.1 mg/kg盐酸丁丙诺啡达到镇痛效果。 小鼠药代动力学和组织分布研究[3] 实验前12小时禁食,但可自由饮水。所有小鼠随机分为两组,分别静脉注射或口服2.6 mg/kg MitoQ。尾部注射(静脉注射)后,分别于0(前药)、2分钟、5分钟、10分钟、15分钟、30分钟、45分钟、1小时、2小时、4小时和6小时采集血样至肝素化试管中。同时,分别于口服给药后0(前药)、2分钟、5分钟、10分钟、15分钟、30分钟、45分钟、1小时、1.5小时、2小时、3小时、4小时和6小时采集血样至肝素化试管中。每个时间点的血样均取自6只小鼠。血样立即在4℃下以3000×g离心10分钟,并将血浆转移至新的1.5 mL Eppendorf管中,然后储存于-80℃直至分析。采用DAS软件(3.0版,中国国家药品监督管理局)的非房室模型方法计算MitoQ的药代动力学参数。绝对生物利用度通过比较口服给药的曲线下面积(AUC)与相同剂量静脉给药的AUC计算得出,方法参照既往报道(Ma et al., 2014)。根据校准曲线得到的MitoQ浓度单位(nmol/L)通过乘以相对分子质量转换为ng/mL。将18只小鼠随机分为三组(每组6只)进行组织分布研究。分别在静脉注射2.6 mg/kg纯MitoQ后10分钟、1小时和2小时处死三组小鼠。随后立即取出睾丸,用生理盐水冲洗,并用滤纸吸干。准确称取0.2 g软组织样本,分别用0.5 ml生理盐水匀浆,并储存于−80 °C直至分析。通过校准曲线得到的MitoQ浓度单位(nmol/L)乘以相对分子质量,转换为ng/mL。 剂量信息/给药方案[3] 动物被随机分为五组(每组n=10)。对照组:小鼠每天依次通过灌胃和腹腔注射给予溶剂(乙醇/0.9%生理盐水=1:9,体积比);TP模型组:小鼠在预先给予上述溶剂后,腹腔注射TP(120 μg/kg);MitoQ组:小鼠分别口服MitoQ(1.3、2.6、5.2 mg/kg),持续1小时,然后每天腹腔注射TP(120 μg/kg)。小鼠经 MitoQ 连续治疗 14 天后,被安乐死,取出睾丸组织并称重,以计算睾丸指数(睾丸重量/体重),并储存在液氮中以备将来研究。 |
| 药代性质 (ADME/PK) |
MitoQ在小鼠体内的药代动力学研究[3]
经验证的LC-MS/MS方法成功应用于小鼠单次口服或尾部注射2.6 mg/kg MitoQ后的药代动力学研究。非房室模型计算的主要药代动力学参数列于图6F。MitoQ的平均血浆浓度-时间曲线如图6D和6E所示。 口服给药后约0.74 h观察到达峰时间(Tmax),表明MitoQ能被快速吸收进入血液循环系统。但血浆峰浓度(Cmax)较低,仅为9.77 ± 2.76 ng/ml。静脉注射和口服给药后,MitoQ 的半衰期 (T1/2) 分别为 1.066 ± 0.319 小时和 0.74 ± 0.49 小时。静脉注射和口服给药后,MitoQ 的绝对生物利用度均较低 (17.95% ± 5.98%)。静脉注射和口服给药后的表观分布容积分别为 76.268 ± 13.534 L/kg 和 324.52 ± 241.44 L/kg。MitoQ 在睾丸组织中的分布情况如图 6G 所示。口服和静脉注射给药后 10 分钟,睾丸组织中 MitoQ 的最高浓度分别为 35.82 ± 8.233 ng/g 和 194.55 ± 8.59 ng/g。然而,在给药2小时后,所有组织样本中均未检测到MitoQ。数据表明,MitoQ未在组织中积累。结果还显示,MitoQ能够穿过血睾屏障,为睾丸损伤提供了物质基础。线粒体醌(MitoQ10甲磺酸盐)是一种靶向线粒体的抗氧化剂,目前正在开发用于治疗神经退行性疾病。本研究旨在建立并验证一种基于液相色谱/串联质谱(LC/MS/MS)的检测方法,用于测定大鼠口服MitoQ10后血浆中的线粒体醌,并检测和鉴定其代谢产物。在进行简单的蛋白质沉淀步骤后,采用反相液相色谱法,以乙腈/水/甲酸为流动相进行梯度洗脱,分析血浆样本。采用正离子模式电喷雾电离多反应监测(MRM)法,以氘代化合物(d3-MitoQ10甲磺酸盐)为内标,分析线粒体醌。线粒体醌的校准曲线在0.5-250 ng/mL浓度范围内呈线性关系,相关系数>0.995。该方法灵敏度高(定量限为0.5 ng/mL),准确度(相对误差<8.7%)和精密度(日内和日间变异系数<12.4%)均可接受。线粒体醌在1.5、20和200 ng/mL浓度下的回收率在87-114%之间。该方法已成功应用于大鼠单次口服给药后的药代动力学研究,初步鉴定出 MitoQ10 的四种代谢物,分别为羟基化 MitoQ10、去甲基 MitoQ10 以及 MitoQ10 醌式葡萄糖醛酸苷和硫酸盐结合物。[4] |
| 参考文献 |
|
| 其他信息 |
用于移植的肾脏大多来自已故捐献者。这些肾脏在移植前必须进行冷保存/储存,以保持组织质量,并为受体选择和运输留出时间。然而,冷保存(CS)可能导致组织损伤、肾脏废弃或移植后长期肾功能障碍。我们之前已确定线粒体超氧化物和其他下游氧化剂是重要的信号分子,它们会导致大鼠肾近端小管细胞的冷保存加复温(RW)损伤。因此,本研究旨在确定在威斯康星大学(UW)保存液中添加线粒体靶向抗氧化剂线粒体醌(MitoQ)是否能够保护肾脏免受冷保存损伤。通过将肾细胞或分离的大鼠肾脏置于单独的UW溶液(4°C,4小时)或含有MitoQ或其对照化合物癸基三苯基溴化膦(DecylTPP)(体外1 μM;离体100 μM)的UW溶液中来诱导冷保存。本研究评估了香烟烟雾暴露后氧化剂生成、线粒体功能、细胞活力和肾脏形态的变化。香烟烟雾导致线粒体超氧化物生成和酪氨酸硝化增加2至3倍,线粒体复合物部分失活,以及细胞死亡和/或肾脏损伤显著增加。MitoQ治疗使氧化剂生成减少约2倍,完全阻止了线粒体功能障碍,并显著改善了细胞活力和/或肾脏形态,而癸基三磷酸酯(DecylTPP)治疗则未提供任何保护作用。这些发现表明,MitoQ可能具有潜在的治疗用途,用于减少器官保存损伤和肾脏丢弃,和/或可能改善肾移植后的肾功能。[1] 总之,这是首篇报道早期香烟烟雾暴露期间线粒体超氧化物显著增加并导致线粒体和肾脏损伤的研究。我们发现线粒体靶向抗氧化剂MitoQ能够显著保护肾近端小管细胞和离体大鼠肾脏免受香烟烟雾(CS)介导的氧化应激、线粒体功能障碍、细胞死亡和肾损伤。这些发现表明,在肾移植前向肾脏输注MitoQ可能具有治疗价值,可以减少CS损伤,改善移植受者的预后,并增加可用于移植的捐献器官数量,所有这些都可能降低医疗保健成本。[1]
尽管氧化应激与急性胰腺炎(AP)的发生密切相关,但迄今为止,抗氧化剂治疗在患者中的疗效并不理想。本研究旨在利用体外和体内方法评估线粒体靶向抗氧化剂MitoQ在实验性AP中的潜在保护作用。MitoQ能够阻断小鼠胰腺腺泡细胞中H2O2诱导的细胞内活性氧(ROS)反应,而对照类似物dTPP则不具备这种作用。 MitoQ 并未降低胆囊收缩素 (CCK) 或胆汁酸 TLCS 诱导的线粒体去极化,且在 10 µM 浓度下本身即可引起去极化。在酶标仪检测中,MitoQ 和 dTPP 均增加了基础细胞死亡和 CCK 诱导的细胞死亡。在 TLCS 诱导的急性胰腺炎 (AP) 模型中,MitoQ 治疗不具有保护作用。在胰泌素过度刺激诱导的 AP (CER-AP) 中,MitoQ 表现出混合效应。观察到组织病理学评分部分改善,这与 dTPP 的作用相似,但未降低胰蛋白酶或血清淀粉酶等生化标志物的水平。有趣的是,在 CER-AP 模型中,MitoQ 同时增加了肺髓过氧化物酶和白细胞介素-6 的水平。MitoQ 对分离的多形核白细胞中活性氧 (ROS) 的产生具有双相效应,抑制了 ROS 的急性升高,但提高了其后期水平。我们的研究结果表明,MitoQ不适用于AP治疗,这与之前对该疾病的抗氧化剂评估结果一致。[2] 总之,本研究结果进一步强调了抗氧化剂疗法不适用于AP治疗,此前一项随机、双盲、安慰剂对照临床试验也证实了这一点。MitoQ对实验性TLCS-AP模型没有保护作用,并且在较轻的CER-AP模型中观察到混合效应,包括炎症标志物升高。这些结果与之前的研究一致,这些研究表明,抑制ROS会通过抑制一种保护性的细胞凋亡机制来增强胰腺腺泡细胞坏死,这种作用会促进AP中的局部胰腺损伤。[2] 总之,本研究证实了MitoQ对TP诱导的体内睾丸损伤具有新的有益作用。在TP诱导的睾丸损伤小鼠模型中,MitoQ有效恢复了睾丸组织的微观结构,恢复了血睾屏障的完整性,并维持了精子发生。这些作用机制可能涉及从两个方面保护睾丸组织。一方面,MitoQ 通过调节线粒体动力学发挥抗氧化作用。另一方面,MitoQ 通过激活 Nrf2/Keap1 信号通路降低氧化应激。其效力、疗效和药代动力学特性使其成为治疗线粒体相关睾丸损伤和男性不育症的潜在临床应用候选药物。[3] |
| 分子式 |
C38H47O7PS
|
|---|---|
| 分子量 |
678.8144
|
| 精确质量 |
678.278
|
| 元素分析 |
C, 67.24; H, 6.98; O, 16.50; P, 4.56; S, 4.72
|
| CAS号 |
845959-50-4
|
| 相关CAS号 |
845959-50-4 (mesylate);845959-52-6 (beta-CD complex);444890-41-9 (cation);336184-91-9 (bromide);
|
| PubChem CID |
11388331
|
| 外观&性状 |
Brown to orange ointment
|
| LogP |
7.706
|
| tPSA |
131.77
|
| 氢键供体(HBD)数目 |
0
|
| 氢键受体(HBA)数目 |
7
|
| 可旋转键数目(RBC) |
16
|
| 重原子数目 |
47
|
| 分子复杂度/Complexity |
965
|
| 定义原子立体中心数目 |
0
|
| SMILES |
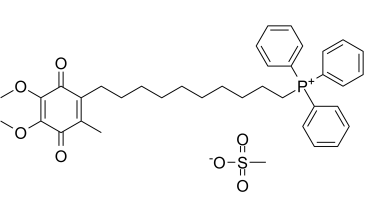
O=C1C(C)=C(CCCCCCCCCC[P+](C2C=CC=CC=2)(C2C=CC=CC=2)C2C=CC=CC=2)C(=O)C(OC)=C1OC.O=S(C)([O-])=O
|
| InChi Key |
GVZFUVXPTPGOQT-UHFFFAOYSA-M
|
| InChi Code |
InChI=1S/C37H44O4P.CH4O3S/c1-29-33(35(39)37(41-3)36(40-2)34(29)38)27-19-8-6-4-5-7-9-20-28-42(30-21-13-10-14-22-30,31-23-15-11-16-24-31)32-25-17-12-18-26-32;1-5(2,3)4/h10-18,21-26H,4-9,19-20,27-28H2,1-3H3;1H3,(H,2,3,4)/q+1;/p-1
|
| 化学名 |
(10-(4,5-dimethoxy-2-methyl-3,6-dioxocyclohexa-1,4-dien-1-yl)decyl)triphenylphosphonium methanesulfonate
|
| 别名 |
Mito Q10; Mitoquinone methanesulfonate; MitoQ; MitoQ10 mesylate; MitoQ mesylate; MitoQ10; MitoQ-10; Mitoquinone mesylate; 845959-50-4; MitoQ; UNII-6E01CG547T; 6E01CG547T; Mitoquinone mesilate; Phosphonium, (10-(4,5-dimethoxy-2-methyl-3,6-dioxo-1,4-cyclohexadien-1-yl)decyl)triphenyl-, methanesulfonate (1:1);
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month 注意: 请将本产品存放在密封且受保护的环境中,避免吸湿/受潮。 |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO : ~100 mg/mL (~147.32 mM)
|
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.5 mg/mL (3.68 mM) (饱和度未知) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 25.0 mg/mL澄清DMSO储备液加入到400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: 2.5 mg/mL (3.68 mM) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (这些助溶剂从左到右依次添加,逐一添加), 悬浊液; 超声助溶。 例如,若需制备1 mL的工作液,可将 100 μL 25.0 mg/mL澄清DMSO储备液加入900 μL 20% SBE-β-CD生理盐水溶液中,混匀。 *20% SBE-β-CD 生理盐水溶液的制备(4°C,1 周):将 2 g SBE-β-CD 溶解于 10 mL 生理盐水中,得到澄清溶液。 View More
配方 3 中的溶解度: 7.69 mg/mL (11.33 mM) in PBS (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液; 超声助溶 (<60°C). 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 1.4732 mL | 7.3658 mL | 14.7317 mL | |
| 5 mM | 0.2946 mL | 1.4732 mL | 2.9463 mL | |
| 10 mM | 0.1473 mL | 0.7366 mL | 1.4732 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
 Effect of MitoQ on mitochondrial superoxide generation during CS.
Effect of MitoQ on CS-induced renal injury and cell death.J Pharmacol Exp Ther.2011 Mar;336(3):682-92. |
|---|
 MitoQ attenuates nitrotyrosine formation during CS.
MitoQ decreases CS plus RW oxidant production and cell death.J Pharmacol Exp Ther.2011 Mar;336(3):682-92. |
 MitoQ prevents mitochondrial respiratory complex inactivation during CS.Individual mitochondrial respiratory complex activities were measured using isolated mitochondria from NRK cells (A and B) and isolated rat kidney mitochondria (C-F) exposed to CS, CS + MitoQ, or CS + DecylTPP (1 μM in vitro and 100 μM ex vivo). Values are expressed as percentage mean ± S.E.M. (n= 3 in vitro or 5 ex vivo) of respective controls (set to 100). **,P< 0.01 or ***,P< 0.001 compared with respective controls. †,P< 0.05 compared with CS.J Pharmacol Exp Ther.2011 Mar;336(3):682-92. |
 HX-1171(HTHQ)
HX-1171(HTHQ)
 新橙皮苷
新橙皮苷
 辛伐他汀羟酸铵盐
辛伐他汀羟酸铵盐
 Disodium (R)-2-Hydroxyglutarate
Disodium (R)-2-Hydroxyglutarate
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1
































 COA
COA


 463611831
463611831
