| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 10 mM * 1 mL in DMSO |
|
||
| 1mg |
|
||
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| 500mg |
|
||
| Other Sizes |
|

| 靶点 |
OST/oligosaccharyltransferase
Oligosaccharyltransferase (OST) complex, including catalytic subunits STT3A and STT3B [1] Oligosaccharyltransferase (OST) complex [2] Oligosaccharyltransferase (OST) complex [3] |
|---|---|
| 体外研究 (In Vitro) |
体外活性:ML414(也称为 NGI-1)是一种新型的细胞渗透性寡糖转移酶 (OST) 抑制剂,OST 是一种异寡聚酶,存在于多种亚型中,可将寡糖转移至受体蛋白。 ML414 是通过基于细胞的高通量筛选和先导化合物优化活动鉴定出来的。在非小细胞肺癌细胞中,NGI-1 阻断表皮生长因子受体 (EGFR) 糖蛋白的细胞表面定位和信号传导,但仅选择性地阻止依赖 EGFR(或成纤维细胞生长因子)的细胞系的增殖,FGFR)以求生存。在这些细胞系中,OST 抑制会导致细胞周期停滞,并伴有 p21 的诱导、自发荧光和细胞形态变化,这些都是衰老的标志。这些结果表明 OST 抑制是治疗受体酪氨酸激酶依赖性肿瘤的潜在治疗方法,并为可逆调节哺乳动物细胞中的 N 连接糖基化提供了化学探针。激酶测定:之前已描述过在 D54-ERLucT 和 D54-LucT 细胞中使用生物发光 N 连接糖基化报告基因的 HTS 方法。简而言之,基于细胞的初级筛选使用具有三个 N 连接糖基化共有序列的修饰和 ER 翻译的荧光素酶蛋白来检测 N 连接聚糖位点占用。与对照相比,D54-ERLucT 中糖基化的抑制可恢复并增加荧光素酶活性,但不会增加非 ER 翻译的 D54-LucT 细胞系中的活性。初级 (D54-ERlucT)、二级假阳性 (D54-LucT) 和三级(荧光素酶抑制)筛选的方法以及使用 CellTitre Glo 进行的毒性测定均存放在 Pubchem (AID 588693) 中。使用具有 Smartfit 算法的 Genedata Screener 软件生成 AC40 值,用于类似物的比较分析。细胞检测:在非小细胞肺癌细胞中,NGI-1 阻断表皮生长因子受体 (EGFR) 糖蛋白的细胞表面定位和信号传导,但仅选择性地阻止依赖于 EGFR(或成纤维细胞生长因子(FGFR)以维持生存。在这些细胞系中,OST 抑制会导致细胞周期停滞,并伴有 p21 的诱导、自发荧光和细胞形态变化,这些都是衰老的标志。这些结果表明 OST 抑制是治疗受体酪氨酸激酶依赖性肿瘤的潜在治疗方法,并为可逆调节哺乳动物细胞中的 N 连接糖基化提供了化学探针。
对拉沙病毒(LASV)及沙粒病毒的作用:有效降低重组沙粒病毒rLCMV/LASV GPC的感染性,且不影响细胞活力;导致LASV糖蛋白(GP)低糖基化,抑制病毒增殖。该作用对新旧世界沙粒病毒具有保守性,在HEK293T、A549、HeLa、Huh7、HEK293细胞中均表现出一致的抗病毒效果[1] - 对NSCLC细胞RTK信号的抑制作用:阻断表皮生长因子受体(EGFR)的细胞表面定位和信号传导;选择性抑制依赖EGFR或成纤维细胞生长因子受体(FGFR)的非小细胞肺癌(NSCLC)细胞(PC9、H1581、H2444)增殖,诱导G1期细胞周期阻滞、p21表达、自荧光产生及衰老相关形态改变;对AR阴性Hela细胞或非RTK依赖的NSCLC细胞(A549)抑制作用微弱[2] - 对胶质瘤细胞RTK活性及放射敏感性的调控作用:在ErbB家族激活水平较高的胶质瘤细胞系(D54、SKMG3、U251、T98G、42-MG)中,减少多种受体酪氨酸激酶(RTK,包括EGFR、ErbB2、ErbB3、MET、PDGFR、FGFR1)的糖基化、蛋白水平及激活;增强细胞对放射治疗的敏感性和化疗药物(替莫唑胺、依托泊苷)的细胞毒性,增加DNA损伤(γH2AX焦点增多)和G1期阻滞;在RTK激活水平低或PTEN缺失的胶质瘤细胞中无上述效果;糖基化非依赖型CD8-EGFR的表达可逆转这些作用,证实RTK抑制是其核心机制[3] |
| 体内研究 (In Vivo) |
NGI-1在体内通过激活ErbB受体减少胶质母细胞瘤的肿瘤生长。[3]
为了评估ML414(NGI-1)对异种移植物肿瘤生长的影响,我们使用了一种克服该化合物低溶解度的ML414(NGI-1)纳米粒子制剂。首先,使用D54 ERLucT异种移植物测试NGI NP的效果,该异种移植物在抑制糖基化后增加了生物发光。我们发现,在24小时(1.7倍,p=.03;图5A)和48小时(1.7倍数,p=.003;图5B)的时间点,接受ML414(NGI-1)的小鼠明显诱导了生物发光。Tunicmaycin是另一种N-连接糖基化抑制剂,用作阳性对照并诱导生物发光(24小时时为4.2倍(p=.007))。这些结果证实了NGI-1 NP在体内抑制D54肿瘤糖基化的能力。[3] 为了评估ML414(NGI-1)在体内的治疗潜力,我们测试了ML414(NGI-1)NP单独和与辐射联合对D54和SKMG3细胞系胶质瘤肿瘤生长的影响。在这些实验中,小鼠被随机分配到四组中的一组接受治疗:对照NP、对照NP+RT、NGI-1NP和NGI-1 NP+RT。每隔一天输送一次ML414(NGI-1)NP(20mg/kg),共3剂,RT以5剂2Gy的日剂量输送。在D54异种移植物中,单独放疗或放疗+NGI-NP治疗显著延迟了肿瘤生长。与仅用放射治疗的患者相比,在RT中添加NGI NP显著降低了肿瘤生长。在39天时,NGI-1 NP+RT组的中位肿瘤体积为566±200 mm3,而单独RT组为1383±305 mm3(p=.001;图5B)。在SKMG3异种移植物中观察到有利于NGI-1 NP和RT联合治疗的类似结果。在这种细胞系中,当作为单一疗法给药时,辐射和NGI NP都能减少肿瘤生长。NGI-1 NPs+RT的组合显著减少了肿瘤生长。放疗+NGI-1-NP组在第98天的平均肿瘤体积几乎无法检测到。相比之下,空白NP的肿瘤体积(379±38 mm3;p=.001)、辐射(139±27 mm3;p=.001)和NGI-1-NP(151±7 mm3;p>0.05)均显著更大(图5C)。对于两种体内异种移植物实验,没有证据表明NGI-NP治疗的动物体重明显减轻或其他毒性。综上所述,这些结果表明NGI-1+RT的组合可能是治疗胶质母细胞瘤的一种治疗方法。[3] 抑制胶质瘤异种移植瘤生长:裸鼠皮下接种D54或SKMG3胶质瘤细胞后,给予ML414 (NGI-1)纳米颗粒制剂(20mg/kg,静脉注射,隔天一次,共3次)联合分次放疗(总剂量10Gy,每日2Gy)。每周测量两次肿瘤体积,结果显示联合治疗组肿瘤生长较单独放疗组或对照纳米颗粒组显著减慢;治疗期间动物未出现明显体重减轻或其他毒性反应[3] - 体内糖基化抑制验证:接种D54-ERLucT异种移植瘤的小鼠,经ML414 (NGI-1)纳米颗粒(20mg/kg,静脉注射)处理后,24小时和48小时的生物发光信号显著增强(1.7倍),证实其在体内可有效抑制N-糖基化[3] |
| 酶活实验 |
先前已描述过在 D54-ERLucT 和 D54-LucT 细胞中使用生物发光 N 连接糖基化报告基因的 HTS 方法。简而言之,基于细胞的初级筛选使用具有三个 N 连接糖基化共有序列的修饰和 ER 翻译的荧光素酶蛋白来检测 N 连接聚糖位点占用。与对照相比,D54-ERLucT 中糖基化的抑制可恢复并增加荧光素酶活性,但不会增加非 ER 翻译的 D54-LucT 细胞系中的活性。初级 (D54-ERlucT)、二级假阳性 (D54-LucT) 和三级(荧光素酶抑制)筛选的方法以及使用 CellTitre Glo 进行的毒性测定均存放在 Pubchem (AID 588693) 中。使用具有 Smartfit 算法的 Genedata Screener 软件生成 AC40 值,用于类似物的比较分析。
细胞-free OST活性测定:在兔网织红细胞裂解物中加入犬胰腺粗微粒体,加入ML414 (NGI-1)或衣霉素,对Saposin-DDK-His6 mRNA进行60分钟翻译反应,通过³⁵S代谢标记检测翻译产物的糖基化水平,评估OST活性[2] - 细胞热迁移实验(CETSA):293T细胞用100μM ML414 (NGI-1)处理30分钟后进行热处理,通过Western blot检测OST亚基的热稳定性变化,验证药物与OST亚基的结合作用[2] |
| 细胞实验 |
在非小细胞肺癌细胞中,NGI-1 阻断表皮生长因子受体 (EGFR) 糖蛋白的细胞表面定位和信号传导,但仅选择性地阻止依赖 EGFR(或成纤维细胞生长因子)的细胞系的增殖,FGFR)以求生存。在这些细胞系中,OST 抑制会导致细胞周期停滞,并伴有 p21 的诱导、自发荧光和细胞形态变化,这些都是衰老的标志。这些结果表明 OST 抑制是治疗受体酪氨酸激酶依赖性肿瘤的潜在治疗方法,并为可逆调节哺乳动物细胞中的 N 连接糖基化提供了化学探针。
沙粒病毒感染及糖基化测定:HEK293T、A549、HeLa、Huh7、HEK293细胞以MOI=0.01感染rLCMV/LASV GPC病毒,病毒进入后加入不同浓度ML414 (NGI-1)(DMSO为对照),36小时后收集细胞和上清液;通过Western blot分析病毒GP的糖基化模式,采用免疫空斑法检测病毒滴度,qRT-PCR测定病毒RNA拷贝数[1] - NSCLC细胞RTK信号及增殖测定:CHO-Lec15、CHO-Lec35、HeLa、HEK293及NSCLC细胞(PC9、A549、H1581、H2444)用1–10μM ML414 (NGI-1)处理24–72小时;Western blot检测EGFR的糖基化、磷酸化及下游信号分子表达;表面生物素化和共聚焦显微镜分析EGFR定位;MTT法检测细胞增殖;流式细胞术分析细胞周期分布;qRT-PCR定量cyclin D1 mRNA水平[2] - 胶质瘤细胞RTK激活及放射敏感性测定:胶质瘤细胞(D54、SKMG3、U251、T98G、42-MG)用10μM ML414 (NGI-1)预处理48小时后,暴露于放射治疗(0–6Gy)或化疗药物(替莫唑胺10μM、依托泊苷0.1μM);克隆形成实验评估放射敏感性;CellTiter 96实验检测细胞增殖;Western blot检测RTK的糖基化、磷酸化及DNA损伤标志物γH2AX;免疫荧光定量γH2AX焦点数量;流式细胞术分析细胞周期分布[3] |
| 动物实验 |
NGI-1在胶质瘤异种移植模型中的治疗研究:[3]
将1×10⁶个D54和SKMG3细胞皮下注射到裸鼠后肢,建立双侧异种移植瘤模型。注射4天后,将小鼠随机分为4个治疗组。分别给予对照组或NGI-1纳米颗粒(20mg/kg)静脉注射,隔日一次,共3次;同时给予假照射或总剂量10 Gy的照射,总剂量分2 Gy/次,使用Precision X-ray 250-kV正电压X射线机进行照射。每周测量两次肿瘤大小,并根据公式π/6 × (大直径) × (小直径)²计算。 所有实验程序均已获得IACUC和耶鲁大学动物护理与伦理委员会以及癌症研究中动物福利和使用指南的批准。使用可检测NLG抑制的生物发光成像平台评估了NGI-1向胶质瘤异种移植瘤的递送。皮下注射1×10⁷个胶质瘤细胞10天后,对携带可触及肿瘤的小鼠进行治疗,分别给予对照组、NGI-1纳米颗粒(20 mg/kg)或衣霉素(1 mg/kg),并在腹腔注射荧光素(150 mg/kg)后5-30分钟进行成像。对包含每个肿瘤的感兴趣区域(ROI)进行信号强度定量,并通过比较治疗前后24小时和48小时的峰值生物发光活性来计算生物发光的诱导情况。[3] 胶质瘤异种移植瘤治疗试验:将1×10⁶个D54或SKMG3胶质瘤细胞皮下注射到裸鼠后肢。四天后,将小鼠随机分为四组:对照纳米颗粒组、ML414 (NGI-1) 纳米颗粒组(20 mg/kg,静脉注射,隔日一次,共3次)、假照射组或照射组(总剂量10 Gy,每日2 Gy)。每周测量两次肿瘤大小,并使用公式π/6 × (大直径) × (小直径)² [3] 计算肿瘤体积。 - 体内糖基化抑制验证试验:将携带D54-ERLucT异种移植瘤(皮下注射1×10^7个细胞)的小鼠分别接受对照纳米颗粒、ML414 (NGI-1) 纳米颗粒(20 mg/kg,静脉注射)或衣霉素(1 mg/kg,静脉注射)治疗。腹腔注射荧光素(150 mg/kg)后 5-30 分钟,分别在 24 小时和 48 小时定量生物发光信号强度,以确认糖基化抑制情况 [3] |
| 毒性/毒理 (Toxicokinetics/TK) |
体外毒性:在抑制病毒感染的浓度下,ML414 (NGI-1) 对 HEK293T、A549、HeLa、Huh7 和 HEK293 细胞的活力没有影响 [1]
- 体内毒性:用 ML414 (NGI-1) 纳米颗粒(20 mg/kg 静脉注射)治疗的小鼠未出现明显的体重减轻或其他明显的毒副作用 [3] |
| 参考文献 |
|
| 其他信息 |
拉沙病毒(LASV)是人类致命性出血热的病原体。LASV的糖蛋白(GP)介导病毒进入宿主细胞,而宿主因子对GP的正确加工和修饰是病毒复制的先决条件。本研究采用亲和纯化-质谱联用(AP-MS)策略,鉴定出591个与LASV GP相互作用的宿主蛋白。通过基因本体分析对这些蛋白进行功能注释,发现寡糖基转移酶(OST)复合物高度富集。利用CRISPR-Cas9介导的基因敲除技术进行的功能研究表明,OST复合物的两个具有催化活性的亚型STT3A和STT3B对于重组沙粒病毒rLCMV/LASV糖蛋白前体的增殖至关重要,其主要作用机制是通过影响病毒的感染性。敲除STT3B(而非STT3A)会导致LASV GP糖基化不足,表明LASV优先依赖STT3B-OST亚型。此外,STT3B-OST的两个特异性亚基——镁转运蛋白1 (MAGT1)和肿瘤抑制候选基因3 (TUSC3)的双敲除也会导致LASV GP糖基化不足,并影响病毒的增殖。定点诱变分析表明,MAGT1或TUSC3的氧化还原酶CXXC活性位点基序对LASV GP的糖基化至关重要。小分子OST抑制剂ML414 (NGI-1)能够有效降低病毒感染性,且不影响细胞活力。STT3B依赖的GP N-糖基化在其他沙粒病毒属中也保守存在,包括旧世界和新世界沙粒病毒属。我们的研究系统地分析了拉沙病毒(LASV)糖蛋白(GP)与宿主的相互作用,并揭示了STT3B对LASV GP N-糖基化的优先依赖性。重要性:糖蛋白在沙粒病毒的生命周期中发挥着至关重要的作用,它们促进病毒进入宿主细胞并参与病毒出芽过程。糖蛋白的N-糖基化是其正常功能的关键;然而,人们对病毒依赖的宿主因子知之甚少。本研究对LASV GP的相互作用组进行了全面的表征,并进一步发现沙粒病毒GP优先依赖STT3B依赖的N-糖基化,且该过程对病毒的感染性至关重要。STT3B-OST复合物的两个特异性硫氧还蛋白亚基MAGT1和TUSC3被发现是病毒GP N-糖基化所必需的。此外,小分子硫氧还蛋白抑制剂ML414(NGI-1)也对沙粒病毒表现出显著的抑制作用。我们的研究为拉沙病毒糖蛋白(LASV GP)与宿主的相互作用提供了新的见解,并拓展了未来开发拉沙热新型疗法的潜在靶点。[1]
天冬酰胺(N)连接糖基化是一种对糖蛋白折叠、稳定性和细胞定位至关重要的蛋白质修饰。为了鉴定能够抑制该生物合成途径中新靶点的小分子,我们启动了一项基于细胞的高通量筛选和先导化合物优化计划,最终获得了一种细胞渗透性抑制剂ML414 (NGI-1)。ML414 (NGI-1)靶向寡糖基转移酶(OST),OST是一种存在多种亚型的异源寡聚酶,能够将寡糖转移至受体蛋白。在非小细胞肺癌细胞中,ML414 (NGI-1) 可阻断表皮生长因子受体 (EGFR) 糖蛋白的细胞表面定位和信号传导,但仅选择性地抑制依赖 EGFR(或成纤维细胞生长因子,FGFR)生存的细胞系的增殖。在这些细胞系中,OST 抑制会导致细胞周期阻滞,并伴有 p21 的诱导、自发荧光和细胞形态改变,这些都是衰老的标志。这些结果表明,OST 抑制是一种治疗受体酪氨酸激酶依赖性肿瘤的潜在治疗方法,并提供了一种可逆地调节哺乳动物细胞中 N-连接糖基化的化学探针。[2] 平行信号传导会降低靶向受体酪氨酸激酶 (RTK) 疗法在胶质瘤中的疗效。我们假设抑制蛋白质N-连接糖基化(一种内质网共翻译和翻译后修饰,对RTK的成熟和激活至关重要)可能为胶质瘤放射增敏提供一种新的治疗方法。实验设计:我们研究了寡糖基转移酶小分子抑制剂ML414 (NGI-1)对EGFR家族受体MET、PDGFR和FGFR1的影响。我们测定了糖基化状态对肿瘤细胞放射敏感性、化疗诱导的细胞毒性、DNA损伤和细胞周期阻滞的影响,并将其与胶质瘤细胞受体表达谱进行关联分析。我们使用经体内分子成像验证的纳米颗粒制剂,测试了ML414 (NGI-1)对异种移植瘤生长的影响。通过表达糖基化非依赖性 CD8-EGFR 嵌合体,评估了 RTK 信号通路的机制作用。结果:ML414 (NGI-1) 降低了大多数 RTK 的糖基化、蛋白水平和活化。ML414 (NGI-1) 还增强了 ErbB 家族活化升高的胶质瘤细胞的放射敏感性和化疗的细胞毒性作用,但在 RTK 活化水平不高的细胞中则无此作用。ML414 (NGI-1) 的放射增敏作用与 DNA 损伤和 G1 期细胞周期阻滞的增加相关。与对照组相比,采用分次放射治疗联合 ML414 (NGI-1) 治疗胶质瘤异种移植瘤可显著抑制肿瘤生长。 CD8-EGFR 的表达消除了 ML414 (NGI-1) 对 G1 期阻滞、DNA 损伤和细胞放射敏感性的影响,从而确定 RTK 抑制是 ML414 (NGI-1) 作用的主要机制。[3] ML414 (NGI-1) 是一种细胞渗透性小分子寡糖基转移酶 (OST) 抑制剂,OST 可催化蛋白质的 N-连接糖基化。它可逆地调节哺乳动物细胞中的N-连接糖基化,而不会完全抑制OST活性[2] ML414 (NGI-1)靶向OST复合物的两个催化亚基(STT3A和STT3B),破坏依赖N-连接糖基化维持稳定性和定位的糖蛋白(例如,RTK、病毒糖蛋白)的成熟和功能[1][2][3] ML414 (NGI-1)通过靶向对病原体/病毒存活和肿瘤进展至关重要的共同生物合成途径,成为RTK驱动的肿瘤(例如,非小细胞肺癌、胶质瘤)和沙粒病毒感染(例如,拉沙热)的潜在治疗药物[1][2][3] 在胶质瘤细胞中,ML414 (NGI-1)主要通过抑制ErbB家族RTK来增强放射敏感性。信号传导,如糖基化非依赖性 CD8-EGFR 的拯救作用所证明 [3] ML414 (NGI-1) 对 RTK 依赖性细胞具有选择性,对非 RTK 依赖性细胞或正常细胞活力的影响极小,表明其具有良好的治疗窗口 [2][3] |
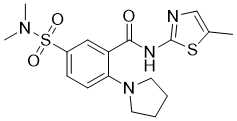
| 分子式 |
C17H22N4O3S2
|
|
|---|---|---|
| 分子量 |
394.51
|
|
| 精确质量 |
394.113
|
|
| 元素分析 |
C, 51.76; H, 5.62; N, 14.20; O, 12.17; S, 16.25
|
|
| CAS号 |
790702-57-7
|
|
| 相关CAS号 |
|
|
| PubChem CID |
2519269
|
|
| 外观&性状 |
White to off-white solid powder
|
|
| 密度 |
1.4±0.1 g/cm3
|
|
| 折射率 |
1.634
|
|
| LogP |
2.75
|
|
| tPSA |
119Ų
|
|
| 氢键供体(HBD)数目 |
1
|
|
| 氢键受体(HBA)数目 |
7
|
|
| 可旋转键数目(RBC) |
5
|
|
| 重原子数目 |
26
|
|
| 分子复杂度/Complexity |
602
|
|
| 定义原子立体中心数目 |
0
|
|
| SMILES |
O=C(C1C(N2CCCC2)=CC=C(S(N(C)C)(=O)=O)C=1)NC1SC(C)=CN=1
|
|
| InChi Key |
QPKGRLIYJGBKJL-UHFFFAOYSA-N
|
|
| InChi Code |
InChI=1S/C17H22N4O3S2/c1-12-11-18-17(25-12)19-16(22)14-10-13(26(23,24)20(2)3)6-7-15(14)21-8-4-5-9-21/h6-7,10-11H,4-5,8-9H2,1-3H3,(H,18,19,22)
|
|
| 化学名 |
5-(dimethylsulfamoyl)-N-(5-methyl-1,3-thiazol-2-yl)-2-(pyrrolidin-1-yl)benzamide
|
|
| 别名 |
NGI-1; NGI1; NGI 1; ML414; 5-(dimethylsulfamoyl)-N-(5-methyl-1,3-thiazol-2-yl)-2-(pyrrolidin-1-yl)benzamide; 5-(N,N-Dimethylsulfamoyl)-N-(5-methylthiazol-2-yl)-2-(pyrrolidin-1-yl)benzamide; MLS002248299; 5-(dimethylsulfamoyl)-N-(5-methyl-1,3-thiazol-2-yl)-2-pyrrolidin-1-ylbenzamide; SMR001315774; ML 414; ML-414;
|
|
| HS Tariff Code |
2934.99.9001
|
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
|
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO : 79~100 mg/mL ( 200.24~253.48 mM )
|
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.5 mg/mL (6.34 mM) (饱和度未知) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 25.0 mg/mL澄清DMSO储备液加入到400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: ≥ 2.5 mg/mL (6.34 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 例如,若需制备1 mL的工作液,可将 100 μL 25.0 mg/mL 澄清 DMSO 储备液加入到 900 μL 玉米油中并混合均匀。 View More
配方 3 中的溶解度: 10% DMSO+40% PEG300+5% Tween-80+45% Saline: ≥ 2.5 mg/mL (6.34 mM); 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.5348 mL | 12.6739 mL | 25.3479 mL | |
| 5 mM | 0.5070 mL | 2.5348 mL | 5.0696 mL | |
| 10 mM | 0.2535 mL | 1.2674 mL | 2.5348 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
|
|
|
|
|
|
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 COA
COA






 463611831
463611831
