| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 10 mM * 1 mL in DMSO |
|
||
| 1mg |
|
||
| 5mg |
|
||
| 10mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| 500mg |
|
||
| Other Sizes |
|

| 靶点 |
Bile acid metabolite; RORγt (Kd = 1.13 μM)
3-Oxo-5β-cholanoic acid targets the transcription factor RORγt, functioning as a direct binder and inhibitor. It binds to RORγt with a Kd of 1.13 μM. RORγt is a key transcription factor that drives the differentiation of TH17 cells, which are pro-inflammatory T helper cells involved in autoimmune and inflammatory diseases. By binding to RORγt, 3-Oxo-5β-cholanoic acid inhibits TH17 cell differentiation. The compound is a metabolite of lithocholic acid, a secondary bile acid. |
|---|---|
| 体外研究 (In Vitro) |
经3-氧代-5β-胆烷酸处理后,RORγt报告基因的活性显著降低。这些数据表明[1],3-氧代-5β-胆烷酸可能通过与RORγt的物理相互作用并抑制其转录活性来抑制TH17细胞的分化。
胆汁酸在哺乳动物肠道中含量丰富,它们在细菌介导下转化,生成大量的生物活性分子。尽管已知胆汁酸会影响宿主代谢、癌症进展和先天免疫,但它们是否会影响表达IL-17a的辅助性T细胞(TH17细胞)或调节性T细胞(Treg细胞)等适应性免疫细胞尚不清楚。我们筛选了一个胆汁酸代谢物库,并鉴定出两种不同的石胆酸 (LCA) 衍生物:3-氧代-5β-胆烷酸/3-氧代石胆酸/3-oxoLCA 和异别石胆酸/isoalloLCA,它们在小鼠体内作为 T 细胞调节剂发挥作用。3-OxoLCA 通过直接结合关键转录因子视黄酸相关孤儿受体-γt (RORγt) 来抑制 TH17 细胞的分化;而 isoalloLCA 则通过产生线粒体活性氧 (mitoROS) 来促进 Treg 细胞的分化,进而导致 FOXP3 表达增加。isoalloLCA 介导的 Treg 细胞分化增强作用依赖于 Foxp3 内含子增强子——保守非编码序列 (CNS) 3;这与先前发现的促进 Treg 细胞分化的代谢物的作用机制不同,后者需要 CNS1。 [1] 3-oxoLCA/3-氧代-5β-胆酸/3-氧代石胆酸抑制Th17细胞分化[1] 接下来,我们检测了3-oxoLCA是否在体外与RORγt蛋白发生物理相互作用。我们使用重组人RORγt配体结合域(LBD)进行了微尺度热泳(MST)实验。结果表明,3-oxoLCA与RORγt LBD存在显著的物理相互作用,其平衡解离常数(Kd)约为1 μM。我们还测试了另外两种结构相似的胆汁酸3-氧代衍生物,即3-氧代胆酸(3-oxoCA)和3-氧代脱氧胆酸(3-oxoDCA)(图2a),结果表明,这两种衍生物的Kd值比3-oxoLCA高约20倍(图2b)。 3-oxoCA 和 3-oxoDCA 对 Th17 细胞分化的抑制作用均不如 3-oxoLCA 显著(图 2c、d)。接下来,我们研究了 3-oxoLCA 是否调节 RORγt 的转录活性。我们检测了胆汁酸对人胚肾 (HEK) 293 细胞中 RORγt 与 Gal4-DBD(DNA 结合域)融合蛋白介导的萤火虫荧光素酶表达的影响14。用 RORγt 特异性拮抗剂 ML209 处理的细胞完全丧失了 RORγt 活性19。同样,3-oxoLCA 处理也显著降低了 RORγt 报告基因的活性(图 2e)。总的来说,这些数据表明 3-oxoLCA 可能通过与 RORγt 发生物理相互作用并抑制其转录活性来抑制 Th17 细胞分化。 3-oxoLCA/3-氧代-5β-胆烷酸/3-氧代石胆酸 对 Th17 细胞的调节作用以及异别石胆酸/异别石胆酸对 Treg 细胞的调节作用具有细胞类型特异性,因为这两种化合物均不影响 T 细胞分化为 Th1 或 Th2 细胞,而分化是通过细胞因子 IFN-γ 和 IL-4 以及转录因子 T-bet 和 GATA3 的表达来评估的(图 1a、b 和扩展数据图 3f、g)。尽管 3-oxoLCA 对 Treg 细胞没有影响(图 1b 和扩展数据图 2e),但 isoalloLCA 可使 Th17 细胞分化减少约 50%,且不影响 RORγt 的表达(图 1a、b 和扩展数据图 3h)。两种化合物均表现出剂量依赖性效应(扩展数据图 4a)。3-oxoLCA 不影响细胞增殖,而与 DMSO 对照组相比,向 T 细胞中添加 isoalloLCA 可导致细胞增殖减少(扩展数据图 4b)。IsoalloLCA 处理不损害细胞活力(扩展数据图 4c)或 T 细胞受体 (TCR) 介导的活化,这由 CD25、CD69、Nur77 和 CD44 等 TCR 活化标志物的相似表达所表明(扩展数据图 4d)。异别胆酸 (isoalloLCA) 可促进 TCR 激活,增强 Treg 细胞功能,因为使用更高浓度的抗 CD3 抗体增强 TCR 激活可显著提高 FoxP3 的表达,且不影响细胞活力(扩展数据图 4e、f)。[1]体外实验表明,3-氧代-5β-胆酸 (3-Oxo-5β-cholanoic acid) 可通过直接结合转录因子 RORγt 抑制 TH17 细胞的分化,其解离常数 (Kd) 为 1.13 μM。该化合物的活性可通过 T 细胞分化实验进行评估,该实验检测 IL-17 和其他 TH17 相关细胞因子的产生。其与 RORγt 的结合可通过表面等离子共振 (SPR) 或等温滴定量热法 (ITC) 等生化实验进行验证。该化合物调节 TH17 细胞分化的能力使其成为研究自身免疫和炎症性疾病机制的宝贵工具。 |
| 体内研究 (In Vivo) |
3-氧代-5β-胆烷酸(脱氢石胆酸)(0.3% (w/w);口服;持续1周)可显著降低回肠中TH17细胞的百分比[1]。
胆汁酸在体内调节T细胞活性[1] 研究人员利用小鼠模型,研究了3-氧代-5β-胆烷酸/3-氧代石胆酸/3-oxoLCA和异别石胆酸/异别石胆酸是否影响体内Th17和Treg细胞的分化。已知节段丝状菌(SFB)是一种小鼠共生菌,可诱导B6小鼠小肠中Th17细胞的分化41。来自Taconic Biosciences (Tac)的C57BL/6NTac小鼠由于SFB的存在,其小肠中含有丰富的Th17细胞。相比之下,来自杰克逊实验室(Jax)的C57BL/6J小鼠缺乏SFB,其肠道Th17细胞数量很少。为了确定3-oxoLCA是否在体内抑制Th17细胞分化,我们用含有SFB的粪便悬液灌胃Jax-B6小鼠,并分别喂食对照饲料或含0.3% (w/w) 3-oxoLCA的饲料,持续一周(图4a)。盲肠内容物中该代谢物的平均浓度为24皮摩尔/毫克湿重(约相当于μM)(扩展数据图7a、b)。该浓度足以在体外抑制Th17细胞分化(图1c)。事实上,3-oxoLCA处理显著降低了回肠Th17细胞的百分比(图4b)。当我们定量分析溃疡性结肠炎患者粪便或常规饲养小鼠盲肠中3-oxoLCA的平均水平时,观察到其平均浓度分别为23和1.0皮摩尔/毫克(扩展数据图7c、d)。对照组和3-oxoLCA处理组的SFB定植水平相当,表明Th17细胞百分比的变化并非由于SFB定植减少所致(扩展数据图7e)。此外,与喂食载体的小鼠相比,喂食3-oxoLCA的Tac-B6小鼠(其体内已存在SFB)的Th17细胞百分比降低(扩展数据图7f-h)。3-oxoLCA处理不影响Treg细胞的百分比(扩展数据图7i)。即使在已知可产生强烈的 Th17 细胞反应的抗 CD3 注射诱导的肠道炎症条件下18,42,经 1% 3-oxoLCA 处理的小鼠(而非 0.3% 3-oxoLCA 处理的小鼠)的 Th17 细胞水平也降低(扩展数据图 7j–l)。 为了研究异别胆酸/异别胆酸 (isoalloLCA) 对体内 Treg 细胞的影响,我们给定植有 SFB 的 B6 小鼠喂食对照饲料或含 0.03% (w/w) isoalloLCA 的饲料。单独使用 isoalloLCA 不足以提高稳态(扩展数据图 7m)和抗 CD3 处理后(扩展数据图 7n)的 Treg 细胞百分比。我们注意到,3-oxoLCA 可进一步增强 isoalloLCA 在体外诱导的 Treg 细胞分化(扩展数据图 7o, p)。与此观察结果一致,0.3% (w/w) 3-氧代-5β-胆烷酸/3-氧代石胆酸/3-氧代LCA 和 0.03% (w/w) 异别LCA 的混合物显著增强了抗CD3治疗小鼠体内的Treg细胞群,与对照组相比(图4c,d)。与体外机制一致,该处理导致回肠固有层CD4+ T细胞线粒体活性氧(mitoROS)生成增加(扩展数据图7q)。重要的是,3-氧代LCA/异别LCA诱导的体内FoxP3表达增强也依赖于CNS3增强子,因为在混合骨髓实验中,ΔCNS3细胞(与野生型细胞不同)不再对该处理产生反应(图4e,f)。在饲料中同时添加异别柠檬酸 (isoalloLCA) 和 3-氧代柠檬酸 (3-oxoLCA) 后,盲肠内容物中异别柠檬酸的平均浓度为 47 皮摩尔/毫克(扩展数据图 7b)。该浓度足以在体外增强 Treg 细胞的分化(图 1c)。人类溃疡性结肠炎患者粪便中异别柠檬酸的平均浓度为 2 皮摩尔/毫克,范围为 0 至 17 皮摩尔/毫克(扩展数据图 7c)。这些数值与喂食 0.03% isoalloLCA 和 0.3% 3-oxoLCA 的小鼠体内观察到的浓度处于同一数量级,表明体内 isoalloLCA 的浓度水平具有生理相关性 [1]。接下来,研究人员探究了 3-氧代-5β-胆烷酸/3-氧代石胆酸/3-oxoLCA 和异别石胆酸/异别石胆酸的免疫调节作用是否是通过改变肠道菌群组成来实现的。对喂食含胆汁酸饮食的小鼠粪便样本进行 16S rDNA 测序,结果显示,与对照组相比,其肠道菌群组成未发生显著变化(扩展数据图 8a–e)。此外,3-oxoLCA 处理降低了感染鼠柠檬酸杆菌的无菌 B6 小鼠结肠中 Th17 细胞的诱导(扩展数据图 8f、g)。因此,3-oxoLCA 和 isoalloLCA 对 Th17 和 Treg 细胞的调节活性可能不需要共生菌群的存在。总之,这些数据表明,3-oxoLCA 和 isoalloLCA 均可在体内直接调节小鼠的 Th17 和 Treg 细胞反应 [1]。 公开文献中关于 3-氧代-5β-胆烷酸的具体体内活性数据尚不充分。作为一种通过 RORγt 结合抑制 Th17 细胞分化的化合物,该化合物在自身免疫性疾病和炎症性疾病的研究中具有潜在的应用价值。可在自身免疫性疾病的动物模型中进行研究,例如实验性自身免疫性脑脊髓炎(EAE)、炎症性肠病或银屑病。还需要进一步的体内研究来表征其疗效、生物利用度和安全性。 |
| 酶活实验 |
哺乳动物荧光素酶报告基因检测[1]
报告基因检测按先前描述的方法进行14。简而言之,将50,000个人类胚胎肾293细胞接种于96孔板中,培养基为不含抗生素的Dulbecco改良Eagle培养基(DMEM),并添加1%胎牛血清(FCS)。将细胞转染DNA混合物,该混合物包含0.5 μg/mL萤火虫荧光素酶报告质粒、2.5 ng/mL含有海肾荧光素酶的质粒以及Gal4-DNA结合域-RORγ(0.2 μg/mL)。转染使用TransIT-293试剂,并按照制造商的说明进行操作。转染24小时后加入胆汁酸或载体对照,16小时后使用双荧光素酶报告基因试剂盒测定荧光素酶活性。 微尺度热泳分析[1] 采用微尺度热泳(MST)分析化合物与RORγ配体结合域(LBD)的结合亲和力。纯化的RORγ-LBD用Monolith NT™蛋白标记试剂盒RED进行标记。将浓度为1 mM至20 nM的系列稀释化合物与55 nM标记的RORγ-LBD在室温下混合,并装入Monolith™标准处理过的毛细管中。使用Monolith NT.115仪器,在20% LED功率和“中等”MST功率下监测热泳,时间设置如下:荧光照射前5秒;MST开启20秒;荧光照射后5秒。使用 NT Analysis 软件(Nano Temper Technologies)拟合 Kd 值。 3-氧代-5β-胆酸的体外酶/受体结合实验通常包括使用 RORγt 蛋白和该化合物进行结合实验。通过剂量反应结合实验评估该化合物与 RORγt 的结合亲和力,并确定其 Kd 值为 1.13 μM。采用表面等离子共振 (SPR)、等温滴定量热法 (ITC) 或荧光偏振等方法进行检测。通过使用 ROR 反应元件-荧光素酶报告基因构建体的功能性分析,证实该化合物能够抑制 RORγt 介导的转录活性。 |
| 细胞实验 |
体外T细胞培养[1]
从指定基因型小鼠的脾脏和淋巴结中,通过FACS分选分离出幼稚CD4+(CD62L+ CD44− CD25− CD4+)T细胞。在某些实验中,使用幼稚CD4+ T细胞分离试剂盒富集幼稚CD4+ T细胞。将40,000个幼稚CD4+ T细胞接种于预先包被仓鼠IgG的96孔板中,并在T细胞培养基(RPMI 1640,10%胎牛血清,25 mM谷氨酰胺,55 µM 2-巯基乙醇,100 U/mL青霉素,100 mg/mL链霉素)中培养,培养基中添加0.25 µg/mL抗CD3抗体(克隆145–2C11)和1 µg/mL抗CD28抗体(克隆37.51)。 Th0细胞培养中,T细胞在培养基中添加100 U/mL的IL-2。Th1细胞分化中,T细胞在培养基中添加100 U/mL的IL-2、10 µg/mL的抗IL-4抗体(克隆11B11)和10 ng/mL的IL-12。Th2细胞分化中,T细胞在培养基中添加10 µg/mL的抗IFNγ抗体(克隆XMG1.2)和10 ng/mL的IL-4。Th17细胞分化中,T细胞在培养基中添加10 ng/mL的IL-6和0.5 ng/mL的TGF-β。Treg细胞培养中,T细胞在培养基中添加100 U/mL的IL-2和不同浓度的TGF-β。大多数体外实验中,为了检测异别LCA的作用,未额外添加TGF-β。胆汁酸、维甲酸、mitoQ或mitoPQ分别在0小时或16小时时间点加入。水溶性低的化合物在加入培养基前进行超声处理。细胞于第3天收集,并通过流式细胞术进行分析。为检测活性氧(ROS)和线粒体膜电位,将培养2天的细胞分别与5 µM mitoSOX、10 µM DCFDA或2 µM JC-1孵育30分钟,然后进行流式细胞术分析。流式细胞术[1] 从体外培养或体内小鼠实验中收集的细胞,在 GolgiPlug 存在下,用 50 ng/mL PMA(佛波醇 12-肉豆蔻酸酯 13-乙酸酯)和 1 µM 离子霉素刺激 4 小时,以测定细胞因子表达。刺激后,用细胞表面标志抗体和 LIVE/DEAD Fixable 染料 Aqua 对细胞进行染色以排除死细胞,然后用 FoxP3/转录因子染色试剂盒进行固定和透化处理,最后用细胞因子和/或转录因子特异性抗体进行染色。所有流式细胞术分析均在 LSR II 流式细胞仪上进行,数据使用 FlowJo 软件进行分析。 细胞增殖实验[1] 将幼稚 CD4+ T 细胞用 1 µM 羧基荧光素琥珀酰亚胺酯标记,并在进行 FACS 分析前培养 3 天。 体外抑制实验[1] 将 2.5 × 104 个来自 CD45.1 B6 小鼠的新鲜纯化的幼稚 CD4+CD25−CD44−CD62Lhigh T 细胞用 1 µM CFSE 标记,在 96 孔圆底板中,用可溶性抗 CD3 (1 µg/mL) 和 5 × 104 个 APC 激活,并在测试细胞 (CD45.2) 存在下培养 3 天。采用流式细胞术评估CD45.1 Tconv细胞的CFSE稀释度。 体外细胞实验利用T细胞培养物评估3-氧代-5β-胆酸对TH17细胞分化的影响。在TH17极化条件下(TGF-β + IL-6),用该化合物处理T细胞。通过ELISA或流式细胞术检测IL-17的产生,并通过qPCR检测RORγt和其他TH17相关基因的表达来评估TH17分化。该化合物抑制TH17分化的能力呈剂量依赖性。采用标准细胞培养条件(37°C,5% CO₂)和合适的培养基。 |
| 动物实验 |
动物/疾病模型: B6 Jax 小鼠(含 SFB 的粪便悬液)[1]
剂量: 0.3% (w/w) 给药途径: 灌胃;持续 1 周 实验结果: 显著降低了回肠 TH17 细胞的百分比。 体内胆汁酸分析[1] 所有胆汁酸的储备液均通过将化合物溶解于分子生物学级 DMSO 中制备。这些溶液用于建立标准曲线。分别使用甘氨胆酸或 β-鼠胆酸 (β-MCA) 作为小鼠和人样品的内标。按照先前报道的方法,从小鼠盲肠和人粪便样品中提取胆汁酸,并使用超高效液相色谱-质谱联用 (UPLC-MS) 进行定量。组织中各胆汁酸的检测限(以皮摩尔/毫克湿重计)如下:β-MCA,0.10;异构体别胆酸(isoalloLCA),0.45;异构体胆酸(isoLCA),0.29;胆酸(LCA),0.12;异构体胆酸(alloLCA),0.43;3-氧代胆酸(3-oxoLCA),0.18。 3-氧代-5β-胆烷酸的体内动物研究通常包括将该化合物给予自身免疫性疾病或炎症性疾病的啮齿动物模型。潜在的研究设计包括多发性硬化症(EAE)、炎症性肠病或银屑病模型。典型的终点包括评估疾病严重程度、测量靶组织中TH17细胞频率和IL-17水平、评估炎症标志物以及药代动力学分析。所有程序必须符合机构动物护理和使用指南。 |
| 药代性质 (ADME/PK) |
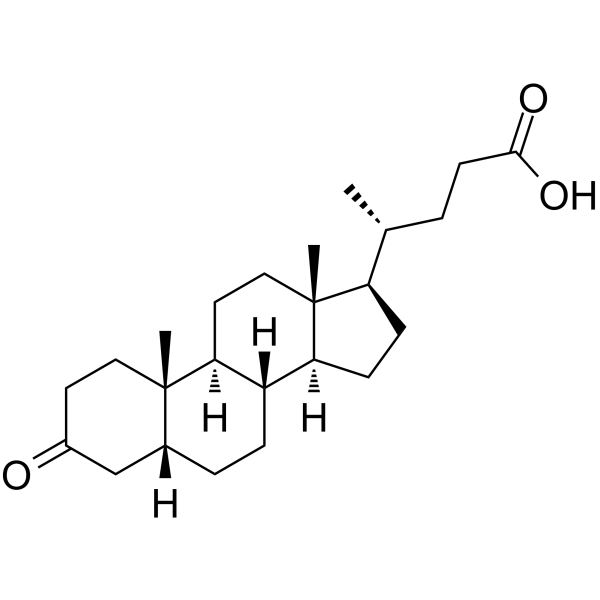
3-氧代-5β-胆酸的分子量为374.56 g/mol,分子式为C₂₄H₃₈O₃。其化学名称为(4R)-4-[(1R,3aS,3bR,5aR,9aS,9bS,11aR)-3a,3b,5a,9b-四氢-9a,11a-二甲基-7-氧代-十六氢-1H-环戊并[a]菲-1-基]戊酸。它是胆汁酸代谢产物,也是石胆酸在人肝脏中的主要CYP3A代谢产物。它通过与RORγt结合抑制TH17细胞分化(Kd = 1.13 μM)。
|
| 毒性/毒理 (Toxicokinetics/TK) |
3-氧代-5β-胆酸仅供研究用途,未获准用于人体治疗。作为一种研究用化学品,公开文献中尚无全面的毒理学数据。其母体化合物石胆酸是一种肝毒性和致癌性胆汁酸。处理该化合物时应遵守标准安全预防措施,包括使用适当的个人防护装备。与所有研究用化学品一样,在考虑进行临床开发之前,必须进行全面的毒理学分析。
|
| 参考文献 | |
| 其他信息 |
3-氧-5β-胆酸是一种氧-5β-胆酸,是3-氧-5β-胆酸盐的共轭酸。一些胆汁酸被认为是组织损伤因子,由于其在肝病患者中的积累增加以及其类似洗涤剂的化学特性,促进炎症。然而,最近的研究开始揭示它们的抗炎作用,特别是在先天免疫系统中,通过抑制NF-κB依赖的信号通路和NLRP3依赖的炎症小体活性。我们的研究揭示了在人体和啮齿动物中发现的两种LCA代谢物的额外抗炎作用,直接影响CD4+ T细胞:3-氧LCA抑制Th17分化,而isoalloLCA促进Treg分化。我们的数据表明,3-氧LCA和isoalloLCA均存在于患有结肠炎的人类粪便样本中,以及在常规喂养的Jax-B6小鼠的盲肠中(扩展数据,图7c,d)。重要的是,这两种胆汁酸在无菌B6小鼠中完全缺失(扩展数据,图7d)。这些数据表明,肠道定植菌可能参与3-氧LCA和isoalloLCA的产生,但我们不能排除宿主酶参与的可能性。考虑到Th17和Treg细胞在多种炎症性疾病中的重要作用及其与肠道定植菌的密切关系,我们的研究建议存在新的调节通路,可以通过胆汁酸代谢物调节T细胞功能。未来对产生3-氧LCA和isoalloLCA的细菌或宿主酶的研究将为在自身免疫疾病和其他炎症性疾病中控制T细胞功能提供新方法。[1]
3-氧-5β-胆酸(去氢胆酸;3-氧LCA)(CAS#: 1553-56-6)的分子式为C₂₄H₃₈O₃,分子量为374.56 g/mol。它是人类肝脏CYP3A代谢的主要胆酸。它通过直接与RORγt结合以Kd 1.13 μM抑制TH17细胞分化。胆酸是一种单羟基次级胆汁酸,是一种肝毒性和致癌性胆汁酸。该化合物不是药物,且未经过临床试验。 |
| 分子式 |
C24H38O3
|
|---|---|
| 分子量 |
374.55672
|
| 精确质量 |
374.282
|
| CAS号 |
1553-56-6
|
| PubChem CID |
5283906
|
| 外观&性状 |
White to off-white solid powder
|
| 密度 |
1.069 g/cm3
|
| 沸点 |
509.3ºC
|
| 闪点 |
275.9ºC
|
| 蒸汽压 |
0mmHg at 25°C
|
| 折射率 |
1.52
|
| LogP |
5.715
|
| tPSA |
54.37
|
| 氢键供体(HBD)数目 |
1
|
| 氢键受体(HBA)数目 |
3
|
| 可旋转键数目(RBC) |
4
|
| 重原子数目 |
27
|
| 分子复杂度/Complexity |
613
|
| 定义原子立体中心数目 |
8
|
| SMILES |
OC(CC[C@H]([C@H]1CC[C@H]2[C@@H]3CCC4CC(CC[C@]4(C)[C@H]3CC[C@]12C)=O)C)=O
|
| InChi Key |
KIQFUORWRVZTHT-OPTMKGCMSA-N
|
| InChi Code |
InChI=1S/C24H38O3/c1-15(4-9-22(26)27)19-7-8-20-18-6-5-16-14-17(25)10-12-23(16,2)21(18)11-13-24(19,20)3/h15-16,18-21H,4-14H2,1-3H3,(H,26,27)/t15-,16-,18+,19-,20+,21+,23+,24-/m1/s1
|
| 化学名 |
(4R)-4-[(1R,3aS,3bR,5aR,9aS,9bS,11aR)-9a,11a-dimethyl-7-oxo-hexadecahydro-1H-cyclopenta[a]phenanthren-1-yl]pentanoic acid
|
| 别名 |
Dehydrolithocholic acid; 3-Oxocholanic acid; 3-oxoLCA; Dehydrolithocholic acid; 1553-56-6; 3-Ketolithocholic acid; 3-Oxo-5beta-cholanoic Acid; 3-keto-lithocholic acid; 3-Oxocholan-24-oic acid; 3-Ketolithocholic acid; 3-Oxolithocholic acid
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month 注意: 请将本产品存放在密封且受保护的环境中(例如氮气保护),避免吸湿/受潮。 |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO: 100 mg/mL (266.98 mM)
H2O: < 0.1 mg/mL |
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.5 mg/mL (6.67 mM) (饱和度未知) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 25.0 mg/mL澄清DMSO储备液加入到400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: ≥ 2.5 mg/mL (6.67 mM) (饱和度未知) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 例如,若需制备1 mL的工作液,可将 100 μL 25.0 mg/mL澄清DMSO储备液加入900 μL 20% SBE-β-CD生理盐水溶液中,混匀。 *20% SBE-β-CD 生理盐水溶液的制备(4°C,1 周):将 2 g SBE-β-CD 溶解于 10 mL 生理盐水中,得到澄清溶液。 View More
配方 3 中的溶解度: ≥ 2.5 mg/mL (6.67 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 配方 4 中的溶解度: 3.33 mg/mL (8.89 mM) in 0.5% CMC-Na/saline water (这些助溶剂从左到右依次添加,逐一添加), 悬浮液; 超声助溶 (<60°C). *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.6698 mL | 13.3490 mL | 26.6980 mL | |
| 5 mM | 0.5340 mL | 2.6698 mL | 5.3396 mL | |
| 10 mM | 0.2670 mL | 1.3349 mL | 2.6698 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 COA
COA
 463611831
463611831
