| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| 500mg |
|
||
| Other Sizes |
|
| 靶点 |
5-HT1A Receptor ( pIC50 = 8.19 ); 5-HT7 Receptor ( pIC50 = 466 nM )
8-OH-DPAT targets human 5-hydroxytryptamine 1A (5-HT1A) receptor with a Ki value of 1.8 nM (radioligand binding assay) [4] 8-OH-DPAT binds to rat 5-HT1A receptor in cerebral cortex with a Ki value of 2.3 nM [1] 8-OH-DPAT shows low affinity for rat 5-HT2 receptors (Ki > 1000 nM) and dopamine D2 receptors (Ki > 500 nM) [1] |
|---|---|
| 体外研究 (In Vitro) |
体外活性:8-OH-DPAT 能够减少培养的人 RPE 细胞中自噬源性和光感受器外节源性脂褐素的积累,增强抗氧化保护并减少氧化损伤。该药物仅对 5-HT1B 亚型较弱有效,pIC50 为 5.42 ± 0.08 (n = 5)。由于 8-OH-DPAT 在浓度低于 100 nM 时对 5-HT1B 结合没有影响。激酶测定:8-OH-DPAT(也称为 8-Hydroxy-DPAT)是一种经典、有效、选择性的 5-HT1A 激动剂,5-HT1A 的 pIC50 为 8.19。它对 5-HT1 结合位点的亚型具有近 1000 倍的选择性,对 5-HT7 的 Ki 为 466 nM; 8-OH-DPAT 与 5-HT1B 弱结合,pIC50 为 5.42,5-HT 的 pIC50 <5。它还能够减少自噬源性和光感受器外节源性脂褐质的积累,增强抗氧化保护并减少培养的人 RPE 细胞的氧化损伤。 5-HT1A受体功能障碍与食欲素KO小鼠的发作性睡病样睡眠功能障碍有关,并且这种功能障碍可能参与了食欲素缺乏引起的睡眠障碍。此外,使用5-HT1A受体激动剂例如8-OH-DPAT可用于治疗食欲素缺乏下的睡眠障碍。细胞测定:将细胞暴露于 H2O2 (200 µM) 中 1 小时,并用 8-OH DPAT (10 µM) 预先或后处理 24 小时。在预处理的情况下,所有测量均在 H2O2 后 24 小时进行,对于后处理,在接触 H2O2 后立即添加 8-OH DPAT。 5HT1A 激动剂处理:为了评估 5-HT1A 受体激动剂 8-OH DPAT 减少培养的 RPE 细胞中脂褐质形成的能力,每 48 小时将该化合物添加到培养基中,浓度范围为 0.1 至 20 µM。所有实验均在基础培养基中进行,接受单独载体(PBS)的细胞作为阴性对照。为了确定 5-HT1A 受体激动剂治疗停止后 8-OH DPAT 的效果是否持续,28 天后停止 8-OH DPAT 治疗,并将细胞维持在基础培养基中或用 POS 喂养另外 28 天。为了评估 8-OH DPAT 去除现有脂褐素的能力,允许自噬衍生的脂褐素和吞噬细胞衍生的脂褐素如上所述累积,然后每隔一天添加 8-OH DPAT,持续长达 28 天。为了确认 8-OH DPAT 通过 5-HT1A 受体激动剂发挥作用,我们在一些实验中加入了 5 µM 5-HT1A 受体拮抗剂 S(-)-UH-301。为了确定 8-OH DPAT 处理时间对氧化应激标记物的影响,将 RPE 培养物用 8-OH DPAT(1 或 10 µM)预处理 3 或 24 小时,然后暴露于 200 µM H2O2 1 小时。小时或暴露于 H2O2 后用 5HT1A 激动剂处理 3 或 24 小时。未暴露于氧化应激源的细胞作为阴性对照,暴露于氧化应激源但未暴露于8-OH DPAT的细胞作为阳性对照。暴露于 8-OH DPAT 的细胞仅充当额外的对照。
在大鼠大脑皮质膜制剂中,8-OH-DPAT 以浓度依赖方式取代[3H]-5-HT与5-HT1A受体的结合,测得Ki值为2.3 nM [1] - 在表达人5-HT1A受体的人胚肾(HEK293)细胞中,8-OH-DPAT 剂量依赖性抑制毛喉素刺激的cAMP积累,EC50为3.6 nM,证实其激动剂活性 [4] - 在原代大鼠皮质神经元中,8-OH-DPAT(10 nM至1 μM)可减轻谷氨酸诱导的细胞毒性,100 nM浓度时细胞存活率提高35%(p < 0.05) [4] - 在表达5-HT1A受体的HEK293细胞中,8-OH-DPAT(1-100 nM)可激活细胞外信号调节激酶(ERK)1/2磷酸化,10 nM时激活程度达对照组的2.8倍 [4] |
| 体内研究 (In Vivo) |
静脉注射选择性 5-HT1A 受体激动剂 8-OH DPAT可迅速逆转严重出血期间产生的低血压和心动过缓反应,且变异性相对较小。 8-OH-DPAT 具有相对亲脂性, 因此可以很容易地穿过血脑屏障。[3]
脑部给药5-羟色胺能5-HT1A受体激动剂延迟对意识大鼠严重出血的反射交感神经溶解反应。为了确定5-HT1A受体激动剂介导这种反应的区域,我们比较了在出血大鼠注射选择性5-HT1A激动剂(+)-8-羟基-2-(二正丙胺)四氢化萘(8-OH DPAT)后,脑室系统或体循环各区域的平均动脉压(MAP)、心率(HR)和肾交感神经活性(RSNA)的恢复情况。三分钟后注入8-OH DPAT(48 nmol /公斤),地图和RSNA高耗费大鼠给药在第四脑室(94 + / - 5毫米汞柱,82 + / - 18%的基线)或体循环(90 + / - 4毫米汞柱,113 + / - 15%的基线)在大鼠给药比外侧渡槽(63 + / - 4毫米汞柱,27 + / - 11%的基线),侧脑室(42 + / - 3毫米汞柱,8 + / - 18%的基线),或者在大鼠给予生理盐水各脑区(47 + / - 5毫米汞柱,-42 + / - 10%的基线)。低剂量注射<强>8-OH DPAT (10 nmol/kg)也能加速出血大鼠的MAP和RSNA的恢复(注射后3分钟为94 +/- 26 mmHg,为基线的72 +/- 33%),但对体循环没有作用(46 +/- 4 mmHg,为基线的-25 +/- 30%)。这些数据表明,8-OH DPAT作用于后脑受体,逆转有意识大鼠出血时的交感神经溶解反应。[3] 5-HT1A激动剂8-OH DPAT介导AMD小鼠模型的神经保护。5-HT1A激动剂8-OH DPAT减少体内脂褐素积累。[4] 在200-250 g雄性Wistar大鼠中,皮下注射8-OH-DPAT(0.1-1 mg/kg)可剂量依赖性降低体温,0.5 mg/kg剂量时最大降温幅度达1.8°C(p < 0.01) [2] - 在C57BL/6小鼠中,腹腔注射8-OH-DPAT(1 mg/kg)可使强迫游泳实验中的不动时间显著减少42%(p < 0.05),提示潜在的抗抑郁样活性 [4] - 在大鼠中,8-OH-DPAT(0.3 mg/kg,皮下注射)可诱导运动减少,与生理盐水对照组相比,旷场实验中的水平活动减少58% [2] - 在自发性高血压大鼠中,静脉注射8-OH-DPAT(0.5 mg/kg)可短暂降低平均动脉血压15 ± 3 mmHg,该效应由外周5-HT1A受体介导 [3] |
| 酶活实验 |
8-OH-DPAT(也称为 8-Hydroxy-DPAT)是一种成熟、强效、特异性的 5-HT1A 激动剂,pIC50 为 8.19。 8-OH-DPAT 与 5-HT1B 弱结合,pIC50 为 5.42,5-HT 的 pIC50 <5;它对 5-HT1 结合位点的亚型具有近千倍的选择性,对 5-HT7 的 Ki 为 466 nM。此外,在培养的人 RPE 细胞中,它可以减少氧化损伤,增强抗氧化防御,并减少自噬和光感受器外节产生的脂褐素的积累。在orexin-KO小鼠中,发作性睡病样睡眠障碍是由5-HT1A受体功能障碍引起的,这也可能导致食欲素缺乏引起的睡眠障碍。此外,使用 5-HT1A 受体激动剂(如 8-OH-DPAT)可能有助于治疗食欲素缺乏引起的睡眠障碍。
5-HT1A受体放射性配体结合实验:将大鼠大脑皮质组织匀浆并离心获得粗制膜组分;膜制剂与[3H]-5-HT及系列浓度的8-OH-DPAT在25°C下孵育60分钟;通过玻璃纤维滤膜快速过滤去除未结合的放射性配体,液体闪烁计数法测定结合放射性;采用非线性回归分析计算Ki值 [1] - cAMP积累实验:将表达人5-HT1A受体的HEK293细胞接种到24孔板,用毛喉素(10 μM)预孵育15分钟;加入8-OH-DPAT(0.1 nM至1 μM)孵育30分钟;裂解细胞后,酶免疫法定量cAMP水平;从剂量-反应曲线推导EC50值 [4] - ERK磷酸化实验:将表达5-HT1A受体的HEK293细胞血清饥饿16小时,用8-OH-DPAT(1 nM至1 μM)处理10分钟;裂解细胞后,蛋白质印迹法检测磷酸化ERK1/2水平;对条带强度进行定量并归一化至总ERK1/2 [4] |
| 细胞实验 |
将细胞暴露于 H2O2 (200 µM) 中 1 小时,并用 8-OH DPAT (10 µM) 预先或后处理 24 小时。预处理阶段的所有测量均在 H2O2 暴露后 24 小时进行,而后处理阶段则立即添加 8-OH DPAT。 5HT1A 激动剂的管理:每 48 小时向培养基中添加 8-OH DPAT(一种 5-HT1A 受体激动剂),浓度范围为 0.1 至 20 µM,以评估该化合物抑制培养干细胞中脂褐质形成的能力。在所有在基础培养基中进行的实验中,仅使用 PBS 的接受细胞作为阴性对照。为了查明 5-HT1A 受体激动剂治疗停止后 8-OH DPAT 的作用是否持续,在 8-OH DPAT 治疗停止后将细胞保留在基础培养基中或用 POS 喂养另外 28 天。为了确定 8-OH DPAT 是否可以消除已经存在的脂褐素,如前所述,允许自噬和吞噬细胞衍生的脂褐素积聚,然后每两天添加 8-OH DPAT,最多 28天。我们在一些实验中加入了 5 µM 5-HT1A 受体拮抗剂 S(-)-UH-301,以验证 8-OH DPAT 通过 5-HT1A 受体激动剂发挥作用。为了确定 8-OH DPAT 处理时间如何影响氧化应激标记物,RPE 培养物在暴露于 H2O2 后用 5HT1A 激动剂处理 3 或 24 小时,或者用 8-OH DPAT 预处理(在 1 或 10 µM) 3 或 24 小时,然后暴露于 200 µM H2O2 1 小时。作为阴性对照,使用未受到氧化应激源的细胞,作为阳性对照,使用受到氧化应激源但不接受 8-OH DPAT 的细胞。暴露于 8-OH DPAT 的细胞的唯一功能是额外的对照。
原代大鼠皮质神经元培养:解剖胚胎18天大鼠的大脑皮质,机械解离后接种到聚L-赖氨酸包被的96孔板,使用神经基底培养基培养;神经元培养7天后进行药物处理 [4] - 谷氨酸诱导的细胞毒性实验:原代皮质神经元用8-OH-DPAT(10 nM至1 μM)预处理1小时,再用谷氨酸(100 μM)处理24小时;基于线粒体脱氢酶活性的比色法评估细胞活力,计算相对于未处理对照组的存活率 [4] - 受体表达验证:将转染人5-HT1A受体cDNA的HEK293细胞在添加胎牛血清的DMEM培养基中培养;转染48小时后,通过蛋白质印迹法和放射性配体结合实验验证受体表达,随后用于功能实验 [4] |
| 动物实验 |
小鼠:每只小鼠的笼内安装一个红外传感器,用于监测动物的运动活动。分别从上午 8:00 和晚上 8:00 开始,每隔 30 分钟记录一次运动活动,以比较光照期和黑暗期的活动情况。所有药物均在晚上 8:00 给药,随后监测 3 小时的运动活动,以确定精神兴奋剂(8-OH-DPAT,1、3 mg/kg,皮下注射等)对黑暗期运动活动的影响。
实验设计 [2] 实验前,将动物连接到记录仪器和抽血泵,并在笼内自由休息。在适应性训练前,将装有脑内注射药物的注射器装满适量液体,并放入引导套管中。然后,让大鼠在出血前至少 2 小时内不受干扰地休息。从出血前10分钟开始,持续记录动脉压、心率和肾交感神经活动(RSNA),直至出血停止后20分钟。控制性抽血以3.2 ml·min⁻¹·kg⁻¹的速率进行6分钟,之后将速率降低至0.53 ml/min,再持续4分钟。初步试验表明,抽取约11.2 ml/kg血液(约占估计血容量的14%)后,平均动脉压(MAP)、心率和肾交感神经活动均出现持续下降。随后降低抽血速度足以维持心动过缓和交感神经阻滞反应,直至出血停止。[2] 开始抽血后(7分钟),将48 nmol/kg的8-OH DPAT(溶于0.5 μl生理盐水)或等体积的生理盐水在20秒内注射到侧脑室、中脑导水管或第四脑室。在第四组中,将药物或溶剂以5 μl的体积静脉注射,并在20秒内用100 μl生理盐水冲洗。所有注射均通过延伸到笼外的导管进行远程操作。药物注射后,在实验结束前未进行任何进一步的干预或复苏。在这些初步研究中,选择的8-OH DPAT剂量与先前实验中确定的ED50相对应,该剂量可增加出血量,并在出血前15分钟注射到侧脑室后导致血压下降40 mmHg。在另一组实验中,以相同方式评估了出血后在第四脑室或全身注射较低剂量(10 nmol/kg)8-OH DPAT的反应。此外,还设置了一个假出血组,该组以相同方式进行操作,并在未进行出血的情况下,在第四脑室注射10 nmol/kg 8-OH-DPAT。[2] 实验结束后,用过量戊巴比妥钠(100 mg/kg,静脉注射)处死大鼠。随后,对插管大鼠进行脑室内注射0.5 μl甲苯胺蓝(0.5%)。迅速取出脑组织,并在10%福尔马林溶液中固定过夜。次日切开脑组织以确认插管位置正确。数据分析仅纳入染料已扩散至脑室系统(即染料未注射到组织中)的动物数据。 8-OH DPAT在SOD2敲低AMD模型中的作用[4] 小鼠被随机分为三组,每日皮下注射无菌生理盐水、0.5 mg/kg体重的8-OH DPAT(溶于无菌生理盐水,低剂量)或5.0 mg/kg体重的8-OH DPAT(高剂量)。每月对小鼠进行一次眼底数字成像、全视野暗适应视网膜电图(ERG)和光谱域光学相干断层扫描(SD-OCT)检查。四个月后,处死小鼠并制备眼球进行冰冻切片。使用共聚焦显微镜测量未经处理的眼睛以及经病毒处理(含或不含8-OH DPAT)的眼睛视网膜色素上皮(RPE)层的自发荧光。激发波长为387 nm,发射波长为440–684 nm,荧光强度使用ImageJ软件进行评估。切片还用于通过光学显微镜和 8-氧代-2′-脱氧鸟苷 (8OHdG) (Abcam,剑桥,马萨诸塞州,美国) 免疫染色评估视网膜结构的损伤,作为氧化应激的测量指标,采用先前描述的方法 大鼠体温和运动研究:雄性 Wistar 大鼠 (200-250 g) 在测试环境中适应 2 小时;8-OH-DPAT 溶解于生理盐水中,并以 0.1、0.3、0.5 或 1 mg/kg 的剂量皮下注射;在给药后 30、60、90 和 120 分钟直肠测量体温;在给药后 30 分钟开始,在开放式场地监测小鼠的运动活性 30 分钟 [2] - 小鼠强迫游泳试验:将 C57BL/6 雄性小鼠(20-25 g)随机分为生理盐水对照组和 8-OH-DPAT 组(每组 n=8);在试验前 30 分钟腹腔注射 8-OH-DPAT(1 mg/kg);将小鼠放入装满水的圆柱体(25°C,20 cm 深)中 6 分钟,并记录最后 4 分钟的静止时间 [4] - 大鼠心血管研究:将自发性高血压大鼠(250-300 g)麻醉,并将导管插入颈动脉以测量平均动脉血压;静脉注射8-OH-DPAT(0.5 mg/kg),并在给药后连续记录60分钟的血压[3] - 初级神经元毒性研究:在胚胎第18天处死妊娠Wistar大鼠,取出胚胎以分离皮质组织用于神经元培养[4] |
| 药代性质 (ADME/PK) |
在大鼠中,静脉注射8-OH-DPAT(1 mg/kg)后,其末端消除半衰期 (t1/2) 为 45 ± 8 分钟 [3]
- 静脉注射8-OH-DPAT后,大鼠体内的分布容积 (Vd) 为 1.2 ± 0.2 L/kg [3] - 大鼠血浆中8-OH-DPAT的血浆蛋白结合率为 28 ± 4%(平衡透析法)[3] |
| 毒性/毒理 (Toxicokinetics/TK) |
在大鼠中,皮下注射8-OH-DPAT(1 mg/kg)可诱发短暂镇静并减少探索行为,这些症状在给药后3小时内消退[2]
- 连续7天皮下注射8-OH-DPAT(0.1-1 mg/kg)的大鼠,其肝脏(ALT、AST)或肾脏(肌酐、BUN)功能指标均未见显著变化[2] - 腹腔注射8-OH-DPAT(最高剂量5 mg/kg)在小鼠给药后24小时内未导致死亡[4] - 皮下注射高剂量8-OH-DPAT(≥2 mg/kg)的大鼠可诱发轻度共济失调和震颤,这些症状是可逆的[2] |
| 参考文献 | |
| 其他信息 |
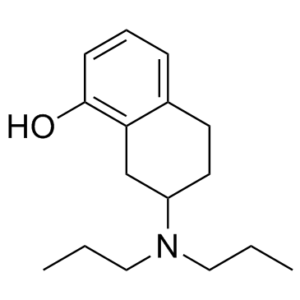
8-OH-DPAT 是一种四氢萘衍生物,其 1 位和 7 位分别被羟基和二丙氨基取代。它是一种血清素能拮抗剂。它属于四氢萘类化合物、酚类化合物和叔胺化合物。它来源于四氢萘的氢化物。
一种血清素 1A 受体激动剂,常用于实验中检测血清素的作用。 两种特异性 5-HT1A 受体激动剂,8-OH-DPAT(0-300 微克/千克)和丁螺环酮(0-3.0 毫克/千克),在变时程阈值电流自刺激大鼠侧下丘脑的实验中进行了测试。丁螺环酮可导致反应的持续单调抑制,而8-OH-DPAT的作用则呈双相性:3.0微克/千克剂量可导致反应的持续增强,而较高剂量(100-300微克/千克)则导致相对短暂的反应抑制。这种双相模式与先前报道的8-OH-DPAT对食物摄入和其他多种行为的影响相一致。阈值电流自刺激对多巴胺能传递的改变高度敏感,但对5-羟色胺(5-HT)的变化相对不敏感。因此,低剂量8-OH-DPAT的促进作用最合理的解释似乎是多巴胺能传递的增强。这可能是由于5-HT1A自身受体介导的5-HT释放抑制以及由此导致的多巴胺能传递去抑制所致。高剂量 8-OH-DPAT 对自我刺激的抑制作用可能反映了 8-OH-DPAT 在突触后 5-HT 受体上的活性,从而抑制了多巴胺 (DA) 的传递。丁螺环酮在所有测试剂量下对反应的抑制作用可能反映了该化合物作为突触后 5-HT 受体部分激动剂的作用,和/或其对其他系统的影响。[2] 年龄相关性黄斑变性 (AMD) 是老年人失明的主要原因之一,与氧化应激、脂褐素积累和视网膜变性有关。本研究旨在确定 5-HT(1A) 受体激动剂是否能在体外和体内减少脂褐素积累、减轻氧化损伤并预防视网膜细胞丢失。本研究采用流式细胞术(FACS)和共聚焦显微镜,在有或无5-HT(1A)受体激动剂8-OH DPAT存在的情况下,评估了培养的视网膜色素上皮(RPE)细胞中自噬来源和光感受器外节(POS)来源的脂褐素的形成。与对照组相比,8-OH DPAT处理导致自噬来源和POS来源的脂褐素均呈剂量依赖性降低。停药后,自噬诱导的脂褐素减少可持续4周。此外,还评估了8-OH DPAT在200 µM H₂O₂处理后减轻氧化损伤的能力。与对照组相比,8-OH DPAT 可降低 H₂O₂ 处理细胞中的超氧化物生成,提高线粒体超氧化物歧化酶 (MnSOD) 水平以及还原型谷胱甘肽与氧化型谷胱甘肽的比值,并能保护细胞免受 H₂O₂ 诱导的脂质过氧化、硝基酪氨酸水平升高和线粒体损伤。具有类似 AMD 表型的 SOD2 基因敲除小鼠每日皮下注射生理盐水、0.5 或 5.0 mg/kg 8-OH DPAT,并每月进行评估。与注射载体的 SOD2 基因敲除小鼠相比,全身注射 8-OH DPAT 可改善 SOD2 基因敲除小鼠眼部的视网膜电图反应。与未治疗的对照组相比,在MnSOD基因敲低后4个月,接受8-OH DPAT治疗的小鼠的ONL厚度显著增加,同时RPE脂褐素减少了60%。这些数据表明,5-HT(1A)受体激动剂可以减少脂褐素的积累,并保护视网膜免受氧化损伤和线粒体功能障碍的影响。 5-HT(1A)受体激动剂可能具有治疗视网膜变性疾病的潜力。[4] 8-OH-DPAT是一种选择性的、中枢活性的5-HT1A受体激动剂,广泛用作血清素受体研究的工具化合物。[1][2][4] - 8-OH-DPAT在皮层神经元中的神经保护作用是通过激活5-HT1A受体、随后的ERK1/2磷酸化以及抑制谷氨酸诱导的兴奋性毒性而介导的。[4] - 8-OH-DPAT诱导的大鼠体温过低依赖于中枢5-HT1A受体的刺激,因为选择性5-HT1A拮抗剂WAY-100635可以阻断这种体温过低。[2] - 8-OH-DPAT调节心血管功能。通过激活外周 5-HT1A 受体,导致血管舒张,从而缓解高血压大鼠的症状[3] - 强迫游泳试验中,8-OH-DPAT 的抗抑郁样活性与增强的血清素能神经传递有关[4] |
| 分子式 |
C16H25NO
|
|
|---|---|---|
| 分子量 |
247.38
|
|
| 精确质量 |
247.194
|
|
| 元素分析 |
C, 77.68; H, 10.19; N, 5.66; O, 6.47
|
|
| CAS号 |
78950-78-4
|
|
| 相关CAS号 |
8-OH-DPAT hydrobromide; 76135-31-4; 8-OH-DPAT;78950-78-4; 141215-27-2 (HCl)
|
|
| PubChem CID |
1220
|
|
| 外观&性状 |
White to gray solid powder
|
|
| 沸点 |
372.5ºC at 760 mmHg
|
|
| 闪点 |
168.2ºC
|
|
| LogP |
3.371
|
|
| tPSA |
23.47
|
|
| 氢键供体(HBD)数目 |
1
|
|
| 氢键受体(HBA)数目 |
2
|
|
| 可旋转键数目(RBC) |
5
|
|
| 重原子数目 |
18
|
|
| 分子复杂度/Complexity |
237
|
|
| 定义原子立体中心数目 |
0
|
|
| SMILES |
OC1C2CC(CCC=2C=CC=1)N(CCC)CCC
|
|
| InChi Key |
ASXGJMSKWNBENU-UHFFFAOYSA-N
|
|
| InChi Code |
InChI=1S/C16H25NO/c1-3-10-17(11-4-2)14-9-8-13-6-5-7-16(18)15(13)12-14/h5-7,14,18H,3-4,8-12H2,1-2H3
|
|
| 化学名 |
7-(dipropylamino)-5,6,7,8-tetrahydronaphthalen-1-ol
|
|
| 别名 |
|
|
| HS Tariff Code |
2934.99.9001
|
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
|
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
|
|||
|---|---|---|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.5 mg/mL (10.11 mM) (饱和度未知) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 25.0 mg/mL澄清DMSO储备液加入到400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: ≥ 2.5 mg/mL (10.11 mM) (饱和度未知) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 例如,若需制备1 mL的工作液,可将 100 μL 25.0 mg/mL澄清DMSO储备液加入900 μL 20% SBE-β-CD生理盐水溶液中,混匀。 *20% SBE-β-CD 生理盐水溶液的制备(4°C,1 周):将 2 g SBE-β-CD 溶解于 10 mL 生理盐水中,得到澄清溶液。 View More
配方 3 中的溶解度: ≥ 2.5 mg/mL (10.11 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 4.0424 mL | 20.2118 mL | 40.4236 mL | |
| 5 mM | 0.8085 mL | 4.0424 mL | 8.0847 mL | |
| 10 mM | 0.4042 mL | 2.0212 mL | 4.0424 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
 |
|---|
 |
 |
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1
































 COA
COA
 463611831
463611831
