| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| 500mg |
|
||
| Other Sizes |
|

| 靶点 |
ML335 and ML402 activate K2P2.1 and K2P10.1 but not K2P4.1, according to two-electrode voltage-clamp studies on Xenopus oocytes (14.3±2.7 μM, K2P2.1-ML335; 13.7±7.0 μM, K2P2.1-ML402; 5.2±0.5 μM, K2P10.1-ML335; and 5.9±1.6 μM, K2P10.1-ML402). K2P2.1 Lys271, which is likewise a lysine in K2P10.1 but a glutamine in K2P4.1, is the only variation between TREK subfamily members at the cation-π interaction site in the K2P modulator pocket. When the Lys271 equivalent is switched between K2P2.1 and K2P4.1, ML335 and M402 activation exhibit a pronounced phenotypic reversal. K2P4.1 (Q258K) responds to both ML402 and ML335 with a smaller magnitude response than K2P2.1[1], but with an EC50 that is similar to K2P2.1 (14.3±2.7 μM, K2P2.1-ML335; 16.2±3.0 μM, K2P4.1(Q258K)-ML335; 13.7±7.0 μM, K2P2.1-ML402; 13.6±1.5 μM, K2P4.1 (Q258K)-ML402)[1].
|
|---|---|
| 体外研究 (In Vitro) |
根据对爪蟾卵母细胞的两电极电压钳研究,ML335 和 ML402 激活 K2P2.1 和 K2P10.1,但不激活 K2P4.1(14.3±2.7 μM,K2P2.1-ML335;13.7±7.0 μM,K2P2.1- ML402;5.2±0.5μM,K2P10.1-ML335;和5.9±1.6μM,K2P10.1-ML402)。 K2P2.1 Lys271 同样是 K2P10.1 中的赖氨酸,但在 K2P4.1 中是谷氨酰胺,是 TREK 亚家族成员之间在 K2P 调节剂口袋中的阳离子-π 相互作用位点处的唯一变异。当 Lys271 等效物在 K2P2.1 和 K2P4.1 之间切换时,ML335 和 M402 激活表现出明显的表型逆转。 K2P4.1 (Q258K) 对 ML402 和 ML335 的响应幅度小于 K2P2.1[1],但 EC50 与 K2P2.1 相似(14.3±2.7 μM,K2P2.1-ML335;16.2± 3.0 μM,K2P4.1(Q258K)-ML335;13.7±7.0 μM,K2P2.1-ML402;13.6±1.5 μM,K2P4.1 (Q258K)-ML402)[1]。
ML402 可激活在非洲爪蟾卵母细胞中表达的 K2P2.1 (TREK-1) 通道,改变其电流-电压关系。该激活作用呈浓度依赖性,EC50 为 13.7 ± 7.0 μM。 ML402 也可激活在爪蟾卵母细胞中表达的 K2P10.1 (TREK-2) 通道,EC50 为 5.9 ± 1.6 μM,但对 K2P4.1 (TRAAK) 通道无作用。 在表达 K2P2.1 (TREK-1) 的 HEK293 细胞的内面向外式膜片中,ML402 (5 μM) 消除了由不对称 K+/Rb+ 条件引起的外向整流,使通道转变为欧姆“泄漏模式”,这表明其直接激活了选择性过滤器的C型门控。在该细胞系统中测得的 EC50 为 5.9 ± 1.6 μM。 ML402 在TREK亚家族(TREK-1, TREK-2)和TRAAK之间的选择性由单个赖氨酸残基(TREK-1中的Lys271)决定。将TREK-1中的该赖氨酸突变为谷氨酰胺(K271Q)会废除对ML402的敏感性,而在TRAAK的等价位置引入赖氨酸(Q258K)则赋予其对ML402的敏感性,且EC50与野生型TREK-1相似。 ML402的激活作用具有特异性,将其芳香族上环替换为脂肪族环的结构类似物未观察到活性,这突出了阳离子-π相互作用对活性的重要性。 其他TREK激活剂如花生四烯酸、BL-1249和ML67-33在Lys271突变体中的作用未改变,表明ML402的作用机制不同。 [1] |
| 细胞实验 |
爪蟾卵母细胞双电极电压钳(TEVC)实验: 向去滤泡的V-VI期非洲爪蟾卵母细胞注射0.15–5 ng编码目标K2P通道的cRNA。24-48小时后,用充满3M KCl的记录电极刺入卵母细胞。在灌流液中,于-80 mV保持电位下记录电流,并通过电压斜坡协议诱发电流。将ML402溶解于DMSO,并在记录缓冲液中连续稀释以进行剂量反应实验。测量并分析电流以确定激活效力(EC50)。
HEK293细胞全细胞膜片钳实验: 用编码K2P2.1 (TREK-1)的质粒转染HEK 293T细胞。转染24小时后,进行全细胞膜片钳记录。测量ML402对0 mV处电流的影响。持续灌流ML402直至达到稳定的增强作用。通过电压斜坡诱发电流。 HEK293细胞内面向外式膜片钳实验: 使用来自转染HEK293细胞的内面向外式膜片,在促进C型门控依赖性整流的条件下(150 mM细胞外K+,150 mM细胞内Rb+)研究电压依赖性激活。在存在和不存在5 μM ML402的情况下,通过电压阶跃诱发电流。计算整流系数以评估向泄漏模式的转变。 [1] |
| 动物实验 |
|
| 参考文献 | |
| 其他信息 |
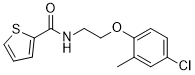
ML402(N-[2-(4-氯-2-甲基苯氧基)乙基]噻吩-2-甲酰胺)是一种噻吩甲酰胺化合物,被鉴定为TREK亚家族双孔域钾离子(K2P)通道的选择性激活剂。
它与位于通道选择性过滤器后方、P1孔螺旋和M4跨膜螺旋交界处的新型“K2P调节口袋”结合。该位点不同于已知的拮抗剂结合位点。 该化合物起到“分子楔”的作用,稳定该界面并降低关键残基(Phe134、Lys271、Trp275)的动态变化。这种稳定作用直接激活了通道的主门控结构——选择性过滤器C型门控结构,从而导致漏电导模式。 TREK-1/TREK-2相对于TRAAK的选择性是由化合物芳香环上部与结合口袋中的赖氨酸残基(TREK-1中的Lys271)之间的阳离子-π相互作用决定的。 本研究揭示了一类新型直接C型门控激活剂的结构基础,并鉴定了一个隐蔽的、可成药的位点,用于调节K2P通道功能。[1] |
| 分子式 |
C14H14CLNO2S
|
|
|---|---|---|
| 分子量 |
295.784461498261
|
|
| 精确质量 |
295.043
|
|
| CAS号 |
298684-44-3
|
|
| 相关CAS号 |
|
|
| PubChem CID |
592973
|
|
| 外观&性状 |
White to off-white solid powder
|
|
| LogP |
3.9
|
|
| tPSA |
66.6
|
|
| 氢键供体(HBD)数目 |
1
|
|
| 氢键受体(HBA)数目 |
3
|
|
| 可旋转键数目(RBC) |
5
|
|
| 重原子数目 |
19
|
|
| 分子复杂度/Complexity |
303
|
|
| 定义原子立体中心数目 |
0
|
|
| SMILES |
C1(C(NCCOC2=CC=C(Cl)C=C2C)=O)SC=CC=1
|
|
| InChi Key |
RULQUKFOBAPKKR-UHFFFAOYSA-N
|
|
| InChi Code |
InChI=1S/C14H14ClNO2S/c1-10-9-11(15)4-5-12(10)18-7-6-16-14(17)13-3-2-8-19-13/h2-5,8-9H,6-7H2,1H3,(H,16,17)
|
|
| 化学名 |
|
|
| 别名 |
|
|
| HS Tariff Code |
2934.99.9001
|
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
|
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
|
|||
|---|---|---|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.5 mg/mL (8.45 mM) (饱和度未知) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 25.0 mg/mL澄清DMSO储备液加入到400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: ≥ 2.5 mg/mL (8.45 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 例如,若需制备1 mL的工作液,可将 100 μL 25.0 mg/mL 澄清 DMSO 储备液加入到 900 μL 玉米油中并混合均匀。 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 3.3809 mL | 16.9045 mL | 33.8089 mL | |
| 5 mM | 0.6762 mL | 3.3809 mL | 6.7618 mL | |
| 10 mM | 0.3381 mL | 1.6904 mL | 3.3809 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
K2P2.1(TREK-1) structures.Nature.2017 Jul 20;547(7663):364-368 |
K2P2.1(TREK-1) activator interactions.Nature.2017 Jul 20;547(7663):364-368 |
K2P2.1(TREK-1) C-tail and lipid binding sites.Nature.2017 Jul 20;547(7663):364-368 |
K2P2.1(TREK-1) activator function.Nature.2017 Jul 20;547(7663):364-368 |
K2Pactivator responses.Nature.2017 Jul 20;547(7663):364-368 |
K2Pchannel patch clamp recordings.Nature.2017 Jul 20;547(7663):364-368 |
 IDOR-1104-0086
IDOR-1104-0086
 Maxi-K modulator 1
Maxi-K modulator 1
 VU6032735
VU6032735
 VU6080824
VU6080824
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 COA
COA






 463611831
463611831
