| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 10 mM * 1 mL in DMSO |
|
||
| 1mg |
|
||
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| Other Sizes |
|

| 靶点 |
TMEM16A/transmembrane protein 16A
T16Ainh-A01 is an inhibitor of the calcium-activated chloride channel TMEM16A (ANO1) (IC50 ~1.1 µM). It also inhibits the related isoform TMEM16B. [2] |
|---|---|
| 体外研究 (In Vitro) |
在不显着影响 L 型 Ca2+ 电流的情况下,T16Ainh-A01 (0.3-30 μM) 大大降低了 IClCa 的内向和外向成分,并抑制 RUICC 中的 IClCa [1]。在所有电压下,TMEM16A 氯化物电流(由移液器中 275 nM 游离钙引起)几乎完全被 T16Ainh-A01 (10 μM) 抑制,表明存在电压无关的阻断机制 [2]。
在跨上皮氯离子梯度下,T16Ainh-A01 (10 µM) 能完全抑制ATP激活的、表达TMEM16A的FRT细胞中的短路电流。 [2] 在原代培养的人支气管上皮细胞中,T16Ainh-A01 (10 µM) 对毛喉素激活的CFTR氯离子电导影响很小(抑制率 <10%)。 [2] 在FRT细胞中,T16Ainh-A01 (10 µM) 不改变由ATP或离子霉素刺激引起的胞质钙升高。 [2] 在全细胞膜片钳记录中,T16Ainh-A01 (10 µM) 在所有的电压下几乎完全抑制了TMEM16A氯离子电流(由电极液内275 nM游离Ca²⁺诱导),表明其阻断机制不依赖于电压。 [2] 在人唾液腺A253细胞中,T16Ainh-A01 (10 µM) 强烈抑制了特征性的外向整流CaCC电流(由电极液内275 nM游离Ca²⁺诱导)。 [2] 在人肠道T84细胞中,T16Ainh-A01 (10 µM) 对ATP诱导的总的、持续的CaCC电流影响甚微,但选择性抑制(约50%)了激动剂刺激后的初始瞬时峰值电流。在HT-29肠道细胞和用UTP刺激的CF患者原代支气管上皮细胞中也观察到对初始峰值电流的部分抑制。 [2] 在气道上皮细胞中,如果在UTP刺激后较晚的时间点加入T16Ainh-A01,其对CaCC电流的抑制作用会减弱,这支持了其主要作用于瞬时电流成分的结论。 [2] 在用IL-4(10 ng/ml,处理24小时)上调TMEM16A表达的人支气管上皮细胞中,UTP诱导的初始峰值电流和T16Ainh-A01敏感的电流成分显著增加。 [2] 在T84细胞中,siRNA敲低TMEM16A在表型上模拟了T16Ainh-A01的作用,减少了早期的瞬时电流,但不影响持续的CaCC电流。 [2] |
| 体内研究 (In Vivo) |
从兔尿道分离的Cajal间质细胞(ICC)表现出Ca2+激活的Cl-电流(I ClCa),这对尿道张力的发展很重要。在这里,我们通过检查“新一代”TMEM16A抑制剂CACCinh-A01和T16Ainh-A01对从新鲜分离的兔尿道ICC(RUICC)记录的I ClCa和对完整兔尿道平滑肌条收缩的影响,来检查TMEM16A(ANO1)是否对此活动有贡献。实时定量PCR实验表明,与TMEM16B和TMEM16F相比,TMEM16A在兔尿道平滑肌中高度表达。单细胞RT-PCR实验表明,新鲜分离的RUICC中仅表达TMEM16A。CACCinh-A01和T16Ainh-A01抑制了用电压钳记录的离体RUICC中去极化诱发的I ClCa,IC50值分别为1.2和3.4μM。同样,CACCinh-A01和T16Ainh-A01也抑制了从-60 mV箝位的RUICC电压中记录的自发瞬态内向电流(STIC)和电流钳中记录的自发性瞬态去极化(STD)。相比之下,CACCinh-A01和T16Ainh-A01仅部分降低了孤立的RUICC中的自发Ca2+波。最后,CACCinh-A01和T16Ainh-A01也显著减少了电场刺激(EFS)引起的兔尿道平滑肌条(RUSM)的神经源性收缩。这些数据与TMEM16A参与RUICC和兔尿道平滑肌收缩的CACCs的观点一致[1]。
|
| 酶活实验 |
筛选过程[2]
如所述,使用配备FluoStar荧光板阅读器的自动筛选平台(Beckman)进行高通量筛选。96孔板的每个孔用PBS(200μl/次洗涤)洗涤三次,留下50μl PBS。将试验化合物(0.5μl)以25μm的终浓度加入每个孔中。10分钟后,将96孔板转移到读板器上进行荧光测定。通过连续记录荧光(400 ms/点)2 s(基线),分别测定每个孔的TMEM16A介导的I-内流,然后加入50μl含有200μm ATP的140 mm I-溶液。I−流入的初始速率是通过非线性回归从荧光数据中计算出来的。[2] 短路电流[2] 将含有表达TMEM16A的FRT细胞、T84细胞或人支气管上皮细胞的Snapwell插入物安装在Ussing室中。将阿米洛利、CFTRinh-172、UTP、ATP和TMEM16A抑制剂加入根尖溶液中,同时将等体积的载体加入基底外侧溶液中。对称的HCO3-缓冲溶液用于T84细胞和人支气管上皮细胞。对于FRT细胞,半腔充满了半Cl-溶液(顶端)和HCO3-缓冲溶液(基底外侧)。将细胞浸泡10分钟稳定期,并在37°C或室温下用95%O2/5%CO2充气。在一些实验中,为了测量根尖氯离子电导率,用制霉菌素(360μg/ml)渗透基底侧膜,并施加氯离子梯度,其中基底侧膜用HCO3-缓冲溶液浸泡,在根尖溶液中用葡萄糖酸钠代替120mm NaCl。使用EVC4000多通道V/I箝位器(佛罗里达州萨拉索塔市世界精密仪器公司)测量短路电流,并使用PowerLab/8sp记录。[2] 贴片夹[2] 在室温下对表达TMEM16A的FRT细胞和人颌下腺A253细胞进行全细胞记录。移液管溶液含有130 mm CsCl、0.5 mm EGTA、1 mm MgCl2、1 mm Tris-ATP和10 mm HEPES(pH 7.2)。浴溶液含有140毫米N-甲基-d-葡糖胺-Cl、1毫米CaCl2、1毫米MgCl2、10毫米葡萄糖和10毫米HEPES(pH 7.4)。移液管由硼硅酸盐玻璃制成,经过火抛光后电阻为3-5兆欧。密封电阻在3到10千欧姆之间。在建立全细胞结构后,TMEM16A被100μm ATP或移液管溶液中275nm游离钙(向移液管液中加入1mm CaCl2)激活。通过施加超极化和去极化电压脉冲,从0 mV的保持电位到-100 mV至+100 mV的电位,以20 mV的步幅,引发全细胞电流。在室温下使用Axopatch-200B进行记录。电流用Digidata 1440A转换器数字化,以5kHz滤波,以1kHz采样。[2] 细胞质钙测量[2] 按照制造商的方案,在镀覆后48小时,将96孔黑壁微孔板中的FRT细胞装载Fluo-4 NW。使用配备注射泵和定制激发/发射滤光片(485/538nm)的FLUOstar Optima荧光板阅读器测量Fluo-4荧光。 |
| 细胞实验 |
全细胞膜片钳记录[1]
采用全细胞膜片钳技术的穿孔贴片配置记录电流。这避免了使用传统全电池配置时遇到的电流耗尽问题。使用抗生素两性霉素B(600μg/ml)穿孔细胞膜。贴片移液管最初通过浸入移液管溶液进行前填充,然后用含两性霉素B的溶液进行后填充。移液管从硼硅酸盐玻璃毛细管(外径1.5 mm,内径1.17 mm)中拔出,尖端直径约为1-1.5μm,电阻为2-4 mΩ。电压钳位命令通过Axopatch 1D膜片钳放大器传递,该放大器连接到与运行pClamp软件的计算机连接的Digidata 1440A AD/DA转换器。在实验过程中,通过一个由移液管(尖端直径200μm)组成的紧密输送系统,将Hanks溶液持续地注入所研究的细胞,该移液管放置在约300μm远的地方。这可以在死区时间约为5秒的情况下切换到含有药物的溶液。所有实验均在35-37°C下进行。[1] 钙显像[1] 将细胞置于含有100μM Ca2+的Hanks溶液中,并让其在玻璃底培养皿中沉淀,直到它们粘在一起。然后将它们在室温下在黑暗中在0.4μM fluo-4/AM中孵育6-8分钟,然后在37°C下进行研究。如前所述,使用iXon 887 EMCCD相机(512×512像素,像素尺寸16×16μm)与Nipkow旋转盘共聚焦头连接对细胞进行成像。使用488nm的氪氩激光器激发fluo-4,在波长>510nm处检测到发射的光。使用×60物镜(奥林巴斯)进行实验,得到像素尺寸为0.266×0.266μm的图像。以每秒15帧的速度采集图像。从每一帧中减去使用零帧获得的相机背景荧光,以获得“F”。F 0被确定为在控制条件下振荡之间测量的最小荧光。为了获得用于在图形中显示的事后行扫描图像,在整个单元格的中心绘制了一条一像素厚的线,并调用了ImageJ中的“reslice”命令。图8a显示了一个例子,其中显示了在自发Ca2+波期间拍摄的RUCC电影堆栈中的选定帧。第一帧中显示的白线表示获得图8b所示伪线扫描的区域。因此,重要的是要认识到,伪线扫描图像仅表示沿线长度发生的事件,而不包括发生在该区域之外的事件。∆F/F 0是指测量Ca2+水平从基础到峰值的变化。 高通量筛选实验: 使用稳定转染了人TMEM16A和卤化物敏感荧光蛋白YFP-H148Q/I152L/F46L的FRT细胞进行TMEM16A抑制剂的筛选。将细胞接种于96孔板,在生理性含氯溶液中与测试化合物(终浓度25 µM)孵育10分钟。然后通过添加含有140 mM I⁻ 和 200 µM ATP的细胞外溶液来触发TMEM16A介导的碘离子内流。通过测量由碘离子内流引起的YFP荧光下降的初始速率来量化抑制效果。 [2] 上皮细胞单层短路电流测量: 将含有细胞单层(表达TMEM16A的FRT细胞、T84细胞或人支气管上皮细胞)的Snapwell小室安装在尤斯灌流室中。细胞浸没在对称的HCO₃⁻缓冲溶液中(对FRT细胞使用半氯溶液),并通以95% O₂/5% CO₂混合气。为了直接测量FRT细胞顶膜TMEM16A电导,使用制霉菌素通透基底膜,并施加跨上皮氯离子梯度。测试化合物添加到顶膜溶液。通过顶膜添加ATP或UTP(100 µM)激活TMEM16A/CaCC。使用电压/电流钳系统测量短路电流。 [2] 全细胞膜片钳记录: 在室温下对表达TMEM16A的FRT细胞或人唾液腺A253细胞进行记录。电极液包含130 mM CsCl, 0.5 mM EGTA, 1 mM MgCl₂, 1 mM Tris-ATP, 10 mM HEPES (pH 7.2),并添加CaCl₂使游离Ca²⁺浓度达到275 nM以激活通道。浴液包含140 mM N-甲基-D-葡糖胺-Cl, 1 mM CaCl₂, 1 mM MgCl₂, 10 mM 葡萄糖, 10 mM HEPES (pH 7.4)。通过从0 mV的保持电位阶跃到-100 mV至+100 mV之间的电压来诱发全细胞电流。通过比较化合物应用前后的电流来评估抑制效果。 [2] 胞质钙测量: 将FRT细胞接种于96孔板并加载Fluo-4 NW染料。使用酶标仪测量荧光。细胞先用化合物或载体预处理5分钟,然后加入钙激动剂(100 µM ATP 或 1 µM 离子霉素)。通过监测荧光变化来评估化合物是否影响钙信号。 [2] 免疫印迹分析: 裂解细胞,蛋白质通过SDS-PAGE分离,转膜,并用TMEM16A一抗进行孵育。使用二抗和增强化学发光法进行检测。此方法用于确认TMEM16A蛋白表达以及siRNA敲低或IL-4处理的效果。 [2] RNA干扰敲低实验: 使用脂质体转染试剂,将靶向TMEM16A的siRNA混合物或非靶向对照siRNA转染到T84细胞中。在24小时重复转染一次。在第二次转染后48小时进行功能实验(短路电流)和免疫印迹分析,以评估TMEM16A敲低的效果。 [2] |
| 动物实验 |
雄性和雌性新西兰白兔(16-20周龄,体重2.5-4 kg)经戊巴比妥钠(静脉注射)安乐死。取其尿道近端1.5 cm,置于克氏液中,并按照先前描述的方法(Sergeant等,2000)[2]酶法分离单个间质细胞(ICC)。
解剖的兔尿道平滑肌条保存在RNAlater中,于-20 °C保存备用。在提取RNA之前,将组织样本转移至1.5 ml离心管中,液氮速冻,研磨成干粉。使用RNeasy Mini试剂盒从这些样本中提取RNA。所有样本均经DNase处理,纯化的RNA用无RNase水洗脱。使用NanoDrop分光光度计测定RNA浓度后,将纯化的RNA保存于-80 °C。在 cDNA 合成之前,RNA 在 70 °C 下变性 5 分钟,然后迅速置于冰上冷却。使用 Superscript VILO cDNA 合成试剂盒,按照制造商的说明进行 RNA 反转录。兔脑组织平行处理作为对照,并按照上述方法从脑组织 RNA 生成 cDNA。[2] 使用 SYBR Green PCR Master Mix 进行实时定量 PCR (qPCR)。设计了 TMEM16B 和 F 基因特异性引物,以覆盖已知 TMEM16 转录本中的外显子-外显子边界。TMEM16A 引物根据本实验室分离的 5′ 端序列设计。在 qPCR 中,β-肌动蛋白用作内源性参考基因进行样本标准化,并据此设计了 β-肌动蛋白引物。[2] |
| 参考文献 |
|
| 其他信息 |
TMEM16A (ANO1) 是一种钙激活氯离子通道 (CaCC)。我们开发了药理学工具,用于研究 TMEM16A 对人呼吸道和肠道上皮细胞中 CaCC 电导的贡献。对约 11 万种化合物的筛选揭示了四类新型小分子 TMEM16A 抑制剂,它们能够完全阻断 TMEM16A 氯离子电流,IC50 < 10 μM,且不干扰钙信号传导。经构效关系分析,其中活性最强的抑制剂为氨基苯基噻唑类化合物 (T16A(inh)-A01),其 IC50 约为 1 μM。我们鉴定了两种不同类型的抑制剂。一些化合物,例如鞣酸和芳基氨基噻吩类化合物 CaCC(inh)-A01,能够完全抑制人支气管和肠道细胞中的 CaCC 电流。其他化合物,包括T16A(inh)-A01和二没食子酸,对这些细胞中的总CaCC电流抑制作用较弱,但主要阻断了激动剂刺激产生的初始瞬时氯离子电流。TMEM16A RNAi敲低也主要抑制了瞬时氯离子电流。与气道和肠道细胞不同,所有TMEM16A抑制剂均能完全阻断唾液腺细胞中的CaCC电流。我们得出结论,TMEM16A几乎承担了唾液腺上皮细胞中所有的CaCC电流,但在气道和肠道上皮细胞中对总CaCC电流的贡献较小。本文鉴定的小分子抑制剂能够对TMEM16A/CaCC的功能进行药理学解析,并有望成为治疗高血压、疼痛、腹泻和黏液分泌过多等疾病的候选药物。[2]
T16Ainh-A01是通过使用TMEM16A转染细胞对约11万个化合物进行基于靶点的高通量筛选而发现的。[2] 它属于2-氨基-4-苯基噻唑衍生物,这类化合物通过硫代乙酰基与第二个杂环(例如嘧啶)连接。构效关系(SAR)分析表明,嘧啶环3、4和5位上的取代基均可耐受,所得类似物的IC₅₀值低至约1 µM。 [2] 该研究得出结论,TMEM16A(可被T16Ainh-A01抑制)是人呼吸道和肠道上皮细胞中总CaCC电导的次要成分,主要负责激动剂刺激后产生的初始瞬时氯离子电流。相比之下,TMEM16A几乎承担了唾液腺上皮细胞中所有的CaCC电流。[2] 用IL-4处理呼吸道上皮细胞可显著上调TMEM16A的表达,并增加T16Ainh-A01敏感的电流成分。[2] 研究结果表明,由于TMEM16A在总呼吸道CaCC电导中的作用较小,因此直接激活TMEM16A可能对囊性纤维化治疗无效,但仍需进行基于表型的筛选来鉴定主要的呼吸道CaCC。[2] |
| 分子式 |
C19H20N4O3S2
|
|---|---|
| 分子量 |
416.5171
|
| 精确质量 |
416.098
|
| 元素分析 |
C, 54.79; H, 4.84; N, 13.45; O, 11.52; S, 15.39
|
| CAS号 |
552309-42-9
|
| PubChem CID |
135460621
|
| 外观&性状 |
White to off-white solid powder
|
| LogP |
4.565
|
| tPSA |
154.26
|
| 氢键供体(HBD)数目 |
2
|
| 氢键受体(HBA)数目 |
7
|
| 可旋转键数目(RBC) |
7
|
| 重原子数目 |
28
|
| 分子复杂度/Complexity |
660
|
| 定义原子立体中心数目 |
0
|
| SMILES |
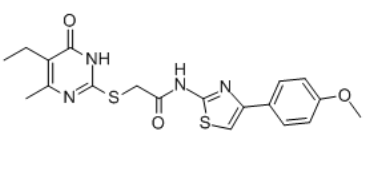
CCC1=C(N=C(SCC(NC2=NC(C3=CC=C(OC)C=C3)=CS2)=O)N=C1O)C
|
| InChi Key |
QSIYTNYMBWYHAA-UHFFFAOYSA-N
|
| InChi Code |
InChI=1S/C19H20N4O3S2/c1-4-14-11(2)20-18(23-17(14)25)28-10-16(24)22-19-21-15(9-27-19)12-5-7-13(26-3)8-6-12/h5-9H,4,10H2,1-3H3,(H,20,23,25)(H,21,22,24)
|
| 化学名 |
2-[(5-ethyl-4-methyl-6-oxo-1H-pyrimidin-2-yl)sulfanyl]-N-[4-(4-methoxyphenyl)-1,3-thiazol-2-yl]acetamide
|
| 别名 |
T16Ainh-A01; T16Ainh A01; T16AInh-A01; 2-((5-Ethyl-4-methyl-6-oxo-1,6-dihydropyrimidin-2-yl)thio)-N-(4-(4-methoxyphenyl)thiazol-2-yl)acetamide; 2-[(5-Ethyl-1,6-dihydro-4-methyl-6-oxo-2-pyrimidinyl)thio]-N-[4-(4-methoxyphenyl)-2-thiazolyl]acetamide; T16Ainh - A01; T16Ainh-A01;; t16a(inh)-a01; 2-[(5-ethyl-6-methyl-4-oxo-1H-pyrimidin-2-yl)sulfanyl]-N-[4-(4-methoxyphenyl)-1,3-thiazol-2-yl]acetamide; T16Ainh-A-01
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO : ~83.33 mg/mL (~200.06 mM)
DMF :≥ 10 mg/mL (~24.01 mM) |
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.08 mg/mL (4.99 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 20.8 mg/mL 澄清 DMSO 储备液加入到 900 μL 玉米油中并混合均匀。 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.4008 mL | 12.0042 mL | 24.0085 mL | |
| 5 mM | 0.4802 mL | 2.4008 mL | 4.8017 mL | |
| 10 mM | 0.2401 mL | 1.2004 mL | 2.4008 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 COA
COA
 463611831
463611831
