| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| 500mg |
|
||
| Other Sizes |
|
| 靶点 |
GABA receptor
|
|---|---|
| 体外研究 (In Vitro) |
在 2/3 层神经元中,THIP(1 µM;5 s)产生强 GABAA 介导的电流 [1]。在第 2/3 层神经元中,Gaboxadol/THIP(1 µM;1 s)对 micro-IPSC 没有影响 [1]。
THIP/Gaboxadol(4,5,6,7-四氢异恶唑[5,4-c]吡啶-3-醇)是一种选择性GABA(a)受体激动剂,偏爱含有GABA(a)受体的δ亚基。THIP目前正在人体试验中测试其催眠效果,显示出有利的耐受性和成瘾性。由于THIP在新皮层中的细胞作用尚不确定,我们研究了THIP对13-19日龄(P13-19)小鼠额顶叶新皮层切片神经元的影响。通过全细胞膜片钳记录,我们发现临床相关的1mum THIP浓度在2/3层神经元中诱导了强劲的GABA(a)介导的强直电流。相比之下,通过模拟体内内源性GABA水平,只诱导了一分钟的紧张电流。微型IPSCs不受1mum THIP的影响,提示其作用位点位于突触外。THIP的EC(50)为44 μ m。THIP诱导的第2/3层神经元的强压电流比第5层神经元的强压电流大44%,与新皮层浅层含有δ受体的表达增强一致。最后,通过监测自发活动的新皮层神经元,THIP引起抑制活性的整体抑制,同时显著增强兴奋性活性。我们的研究表明,THIP激活了突触外GABA(A)受体介导的新皮层传导,这可能会改变皮层网络的活性。[1] Gaboxadol/加博沙多 转运通过Caco-2细胞单层的表征[2] 在不同的加博沙多浓度、顶端pH值以及缺乏或存在5-HTP的情况下,研究了加博沙多在Caco-2细胞单层上的经上皮转运(表1)。在质子梯度穿过单层的情况下(顶端和基底侧腔的pH值分别为6.0和7.4),加博沙多的平均a - b通量增加,而加博沙多的渗透率分别从8.0 × 10 - 6下降到6.1 × 10 - 6和5.6 × 10 - 6 cm s - 1 (p < 0.05),分别为0.34、3.5和7.0 mM加博沙多(表1)。因此,加博沙多在Caco-2细胞中的渗透性是浓度依赖性的,这与hPAT1的部分饱和是一致的。在没有质子梯度的情况下(顶端和基底侧腔的pH值分别为7.4和7.4),3.5 mM和7 mM加博沙多的渗透性分别降低了约5倍和7倍(p < 0.01和p < 0.001)。这与加博沙多通过hPAT1转运是一致的,因为转运体是质子偶联的。在5-HTP存在的情况下,加博沙多的通透性降低了约5倍(p < 0.001)。加博沙多在B-A方向的转运是A-B方向转运的21%,因此加博沙多在A-B方向上的转运是极化的(p < 0.01)。加博沙多在没有质子梯度或有5-HTP存在时的渗透率与加博沙多在B-A方向的渗透率和[14C]-甘露醇的渗透率非常相似。总之,这些观察结果表明,质子依赖的氨基酸转运蛋白hPAT1介导了加博沙多在肠细胞刷状边界膜上的转运。此外,结果表明hPAT1在Caco-2细胞单层中负责80-90%的加博沙多经上皮转运。 盐酸加波沙朵/Gaboxadol(0.34、3.5和7.0 μM)以剂量依赖性方式降低Caco-2单层细胞的通透性,平均Papp值为8.1 × 10-6 cm·s-1、6.1 × 10 -1 cm·s-1(0.34、3.5 和 7 μM 加波沙朵),5.6 × 10-6 cm·s-1(0.34、3.5 和 7 μM 加波沙朵)[6]。 Gaboxadol在体外通过hPAT1转运的特性 [6] 在Gaboxadol/加博沙多浓度增加的情况下,通过测量hPAT1底物脯氨酸进入cco -2细胞单层的顶端摄取,研究了加博沙多和hPAT1之间的相互作用(图1)。加博沙多降低cco -2细胞单层的顶端脯氨酸摄取,估计抑制剂亲和力(Ki值)为6.6 mmol·L−1。同样,已知的PAT1抑制剂色氨酸也降低了脯氨酸的顶端吸收,Ki值为7.7 mmol·L−1。研究了三种浓度(0.34、3.5和7.0 mmol·L−1)下加博沙多在caco2细胞单层上的经上皮转运(A-B)通量。对于0.34、3.5和7 mmol·L−1的加博沙多,加博沙多转运的平均Papp值分别为8.1 × 10−6 cm·s−1、6.1 × 10−6 cm·s−1和5.6 × 10−6 cm·s−1(图2)。因此,随着加博沙多浓度的增加,加博沙多通过Caco-2细胞单层的通透性降低(P < 0.05)。在35 mmol·L−1色氨酸存在的情况下,研究了加博沙多在caco2细胞单层中使用3.5 mmol·L−1加博沙多的顶端浓度,有或没有pH梯度,双向运输(图2)。加博沙多在A-B方向的转运量约为B-A方向的5倍(P < 0.005)。色氨酸的存在使加博沙多的通透性降低了53% (Papp为2.9 × 10−6 cm·s−1,P < 0.005)。在没有质子梯度的情况下,加博沙多的渗透率降低了82%,为1.1 × 10−6 cm·s−1 (P < 0.005)。在色氨酸存在、质子梯度不存在的情况下,加博沙多在B-A方向的渗透性与[3H]-甘露醇的渗透性相似(Papp为1.6±0.36 × 10−6 cm·s−1)。此外,在转运实验中,加博沙多和色氨酸的存在并没有改变美托洛尔或甘露醇在Caco-2细胞单层间转运的通透性。甘露醇和美托洛尔的渗透率分别为1.6±0.36 × 10−6 cm·s−1和6.9±0.99 × 10−6 cm·s−1。综上所述,加博沙多通过Caco-2细胞单层的上皮转运是ph依赖的,可以被色氨酸抑制,并在A-B方向极化。综上所述,这些观察结果表明,hPAT1介导了加博沙多在人肠上皮细胞腔膜上的转运,这一转运步骤在很大程度上决定了加博沙多的上皮转运。 加博沙多/Gaboxadol转运通过hPAT1体外的表征[6] 在加博沙多浓度增加的情况下,通过测量hPAT1底物脯氨酸进入cco -2细胞单层的顶端摄取,研究了加博沙多和hPAT1之间的相互作用(图1)。加博沙多降低cco -2细胞单层的顶端脯氨酸摄取,估计抑制剂亲和力(Ki值)为6.6 mmol·L−1。同样,已知的PAT1抑制剂色氨酸也降低了脯氨酸的顶端吸收,Ki值为7.7 mmol·L−1。研究了三种浓度(0.34、3.5和7.0 mmol·L−1)下加博沙多在caco2细胞单层上的经上皮转运(A-B)通量。对于0.34、3.5和7 mmol·L−1的加博沙多,加博沙多转运的平均Papp值分别为8.1 × 10−6 cm·s−1、6.1 × 10−6 cm·s−1和5.6 × 10−6 cm·s−1(图2)。因此,随着加博沙多浓度的增加,加博沙多通过Caco-2细胞单层的通透性降低(P < 0.05)。在35 mmol·L−1色氨酸存在的情况下,研究了加博沙多在caco2细胞单层中使用3.5 mmol·L−1加博沙多的顶端浓度,有或没有pH梯度,双向运输(图2)。加博沙多在A-B方向的转运量约为B-A方向的5倍(P < 0.005)。色氨酸的存在使加博沙多的通透性降低了53% (Papp为2.9 × 10−6 cm·s−1,P < 0.005)。在没有质子梯度的情况下,加博沙多的渗透率降低了82%,为1.1 × 10−6 cm·s−1 (P < 0.005)。在色氨酸存在、质子梯度不存在的情况下,加博沙多在B-A方向的渗透性与[3H]-甘露醇的渗透性相似(Papp为1.6±0.36 × 10−6 cm·s−1)。此外,在转运实验中,加博沙多和色氨酸的存在并没有改变美托洛尔或甘露醇在Caco-2细胞单层间转运的通透性。甘露醇和美托洛尔的渗透率分别为1.6±0.36 × 10−6 cm·s−1和6.9±0.99 × 10−6 cm·s−1。综上所述,加博沙多通过Caco-2细胞单层的上皮转运是ph依赖的,可以被色氨酸抑制,并在A-B方向极化。综上所述,这些观察结果表明,hPAT1介导了加博沙多在人肠上皮细胞腔膜上的转运,这一转运步骤在很大程度上决定了加博沙多的上皮转运。 Gaboxadol是Caco-2细胞单层中质子偶联氨基酸转运体hPAT1的底物[6] 加博沙多抑制cco -2细胞对hPAT1底物脯氨酸的顶端吸收,Ki值为6.6 mmol·L−1。这种亲和力与最近观察到的其他hPAT1底物如GABA (3.1 mmol·L−1)和GABA类似物muscimol (1.7 mmol·L−1)和THPO (11.3 mmol·L−1)的亲和力相当(Larsen et al., 2008)。色氨酸的亲和力与加博沙多相当,为7.7 mmol·L−1。Metzner等人(2005)先前将色氨酸描述为PAT1的抑制剂,并报道了cco -2细胞中通过hPAT1摄取脯氨酸的Ki值为4.7 mmol·L−1。考虑到两个实验室对hPAT1脯氨酸亲和力的微小差异;Metzner等人报告的Kt值为1.4 mmol·L−1,而Larsen等人报告的Km值分别为3.6 mmol·L−1,两项研究中色氨酸对hPAT1的亲和力相当(Metzner等人,2005;Larsen et al., 2008)。根据其他小组发表的结果(Thwaites et al., 1993;Metzner et al., 2005),我们发现ccao -2细胞的大部分顶端脯氨酸运输是由hPAT1介导的,没有观察到脯氨酸的其他钠依赖性或钠依赖性转运体的证据(Larsen et al., 2008)。其他氨基酸转运蛋白,如顶钠依赖性氨基酸转运蛋白B0 (B0AT1), B0,+ (ATB0,+)和系统(ASC) (ASCT2)不太可能参与加博沙多的转运。它们都存在于Caco-2细胞中,但它们的底物移位不是质子偶联的。此外,与PAT1相比,这些转运蛋白对底物的亲和力值更高,例如ASC (ASCT2),约为100 μ mol·L−1 (Uchiyama et al., 2005);B0 (B0AT1), 500-700µmol·L−1 (Broer et al., 2004)和B0,+ (ATB0,+),约150µmol·L−1 (Hatanaka et al., 2002)。 Gaboxadol在Caco-2细胞单层间的上皮运输呈尖向基底侧极化。色氨酸可以抑制加博沙多的转运,并依赖于顶端供体溶液的pH。此外,随着加博沙多浓度的增加,加博沙多在根尖-基底侧方向的Papp降低。这与加博沙多通过hPAT1转运一致,该途径约占总上皮转运的80%。在小鼠小肠和Caco-2细胞中发现了氨基酸转运系统b0,+,分别占丙氨酸和精氨酸总转运量的15-85% (Wenzel et al., 2001;Dave et al., 2004)。然而,色氨酸与系统b0 +的结合尚未明确显示(Su et al., 1992;Tate et al., 1992),此外,阳离子氨基酸、两性离子氨基酸和胱氨酸对体系b0,+具有µmol·L−1的亲和性(Palacin, 1994)。因此,如果加博沙多通过系统b0,+运输到任何显著程度,它在Caco-2细胞中应该是明显的。在体外,经上皮的加博沙多转运还具有ph依赖性,而PAT1是目前已知的肠道中唯一的质子偶联氨基酸转运蛋白。加巴喷丁(也是两性离子γ-氨基类似物)在大鼠小肠中的渗透性被证明是质子独立的(Nguyen et al., 2007),因此加巴喷丁和加博沙多存在不同的根尖转运机制。结果表明,加博沙多是Caco-2细胞单层中hPAT1的底物,hPAT1介导加博沙多通过肠道肠细胞的管腔膜运输,这似乎对由此产生的上皮运输很重要。加博沙多跨基底外膜外排的机制尚不清楚。 加博沙多/Gaboxadol的体内吸收 在狗[6] 口服给药后,加博沙多在犬体内吸收迅速,Tmax约为0.46 h,生物利用度高达85%。这些观察结果与先前关于人体口服吸收的研究一致,表明加博沙多的Tmax约为0.5 h,生物利用度约为90% (Schultz等人,1981;Lund et al., 2006)。一旦被吸收,加博沙多主要以加博沙多的形式从尿液中排出,而一小部分以葡萄糖醛酸偶联物的形式排出,在大鼠和小鼠中占2-7%,在两个人类受试者中占30-35% (Schultz et al., 1981;Lund et al., 2006;Shadle et al., 2006)。总的来说,这表明在狗体内,加博沙多被迅速和完全吸收,可能在小肠近端,吸收后代谢最小。 |
| 体内研究 (In Vivo) |
加波沙朵(THIP;0.5、5.0 mg/kg;口服;单剂量)表现出良好的口服利用度;在 0.5 和 5.0 mg/kg 时,Fa 值分别为 83% 和 110% [2]。
研究了大鼠口服加博沙多/Gaboxadol在预剂量5-HTP存在和不存在的情况下的体内药代动力学特征。在Caco-2细胞单层>中,80%的吸收性加博沙多转运被认为是hpat1介导的。在大鼠体内,加博沙多的初始吸收率在5-羟色胺的存在下降低。5-羟色胺预给药后,加博沙多的AUC增加3.6 ~ 5.5倍。加博沙多是肾转运蛋白rOat1的底物,K(m)值为151微米。5- http不与rOat1相互作用。综上所述,加博沙多作为hPAT1的底物,也是rOat1的底物。在大鼠中,5-HTP降低了加博沙多的初始吸收率,增加了AUC。因此,5-HTP对加博沙多的药代动力学有显著影响。[2] 盐酸加波沙朵/Gaboxadol(腹膜内注射;0.5、1、1.5、2、3、4 或 5 mg/kg;每天三次;间隔三天)使 Fmr1 KO2 小鼠的步行距离正常化至 0.5 mg/kg WT 活动水平,此外,该化学物质对 Fmr1 KO2 小鼠的运动活性没有影响 [5]。 在这里,我们试图评估Gaboxadol(也称为OV101和THIP),一种选择性和有效的含有δ亚基的突触外GABAA受体(dSEGA)的激动剂,通过评估其在相对未表征的FXS小鼠模型(Fmr1 KO2小鼠)中的异常行为正常化能力,作为FXS的治疗剂。四个行为领域(多动、焦虑、攻击和重复行为)通过一系列行为分析进行了探讨。结果显示,与野生型(WT)相比,Fmr1 KO2小鼠过度活跃,具有异常焦虑样行为,更易怒和更具攻击性,重复行为频率增加,这些都是FXS个体的行为缺陷。加博沙多/Gaboxadol治疗使Fmr1 KO2小鼠中观察到的所有异常行为恢复到WT水平,为其治疗FXS的潜在益处提供了证据。我们发现,仅加博沙多就能增强突触外GABA受体,足以使FXS模型中的许多行为缺陷正常化,这些缺陷的终点可直接转化为FXS的临床表现。综上所述,这些数据支持未来对FXS患者加博沙多的评估,特别是关于多动症、焦虑、易怒、攻击和重复行为的症状。[5] 加博沙多使Fmr1 KO2小鼠的过度活动正常化[5] 多动是人类FXS的一个显著特征(Bailey et al., 2008;Wheeler et al., 2014;Hagerman et al., 2017),并已在先前表征的荷兰-比利时Fmr1 KO小鼠中可靠地复制(Olmos-Serrano et al., 2010;Kazdoba et al., 2014)。为了测试Fmr1 KO2小鼠是否表现出运动亢进,以及加博沙多是否能使这种异常行为正常化,我们给Fmr1 KO2小鼠注射了载药或加博沙多(0.5-5 mg/kg, i.p),并在OFT测试前30分钟给WT窝仔注射载药。记录30 min在OFT内行走的总距离(cm)。结果显示,Fmr1 KO2小鼠的行走距离较WT窝中对照组显著增加(图1,F(8,81) = 21.27, p < 0.0001),与其他FXS模型的结果一致。加博沙多(0.5 mg/kg)使Fmr1 KO2小鼠的行走距离正常化到WT活性水平(图1)。高剂量的加博沙多(1 - 5 mg/kg, ig)对Fmr1 KO2小鼠的运动活性没有影响(图1)。这些结果不能归因于加博沙多的镇静作用,因为在WT C57Bl/6或BALB/c小鼠中,加博沙多高达2.0 mg/kg的剂量在60分钟的OFT中对运动活动没有影响(数据未显示),这与先前的研究一致,表明加博沙多对WT小鼠(Olmos-Serrano等人,2011)或大鼠(Silverman等人,2016)的运动没有影响。 Fmr1 KO2小鼠焦虑样行为经加博沙多归一化 [5] 为了评估加博沙多对Fmr1 KO2小鼠焦虑样行为的影响,采用了三种不同的行为测试:OFT、LDT和SAT的中心移动距离。中心移动距离的增加被解释为焦虑的减少,并利用了小鼠在进入新环境时保持在周边的固有偏好。Fmr1 KO2小鼠注射加博沙多(0.5-5 mg/kg, i.p), WT窝仔在放置于OFT中30分钟前注射载药。与WT对照组相比,Fmr1 KO2小鼠在中心行走的总距离显著增加(图2A, F(8,81) = 21.32, p < 0.0001)。加博沙多治疗(0.5 mg/kg, i.p.p)使Fmr1 KO2对中心距离的影响正常化,达到与WT对照组相当的水平(图2A)。在本实验中,高剂量的加博沙多(1-5 mg/kg)对Fmr1 KO2小鼠没有影响(图2A)。 加博沙多使Fmr1 KO2小鼠的易怒和攻击行为正常化 [5] 与其他形式的综合征自闭症一样,很大一部分FXS患者表现出易怒、社交焦虑和攻击性。这些异常行为可以在啮齿类动物中建模,通过表征测试小鼠和新笼伴侣之间的si。为了验证Fmr1 KO2突变体易怒和攻击性增加的假设,我们量化了摇尾、咬人行为、攀爬行为和攻击延迟的实例。小鼠入笼前30 min分别注射载药或加博沙多(0.5 ~ 5mg /kg, i.p)。 尾巴嘎嘎作响,或尾巴的快速振动,反映了攻击性和战斗倾向。与WT对照组相比,Fmr1 KO2小鼠的摇尾频率显著增加(图3A, F(8,81) = 16.03, p < 0.0001)。加博沙多(0.5、1.5和5.0 mg/kg)使Fmr1 KO2小鼠的效果正常化,达到与WT对照组相当的水平(图3A)。 Gaboxadol使Fmr1 KO2小鼠的重复行为正常化[5] 坚持和重复行为在FXS患者中很常见,并且具有高度破坏性(Arron et al., 2011;Leekam et al., 2011;Hall et al., 2016)。为了验证这些特征可能在Fmr1 KO2动物中观察到的假设,我们量化了WT和Fmr1 KO2突变小鼠的打圈、自我梳理和刻板印象。小鼠分别注射载药或加博沙多(0.5 ~ 5mg /kg, i.p)后,在实验室内测量逆时针转数(CCW)。与WT对照组相比,Fmr1 KO2小鼠在5分钟测试期间CCW转数显著增加(图4A, F(8,81) = 25.46, p < 0.0001)。向Fmr1 KO2小鼠注射加博沙多(0.5、1.0 mg/kg)后,CCW转数恢复到WT水平(图4A)。基因型对顺时针旋转无影响(p = 0.386,数据未显示)。 加博沙多的体内吸收 在狗[6] 口服给药后,加博沙多在犬体内吸收迅速,Tmax约为0.46 h,生物利用度高达85%。这些观察结果与先前关于人体口服吸收的研究一致,表明加博沙多的Tmax约为0.5 h,生物利用度约为90% (Schultz等人,1981;Lund et al., 2006)。一旦被吸收,加博沙多主要以加博沙多的形式从尿液中排出,而一小部分以葡萄糖醛酸偶联物的形式排出,在大鼠和小鼠中占2-7%,在两个人类受试者中占30-35% (Schultz et al., 1981;Lund et al., 2006;Shadle et al., 2006)。总的来说,这表明在狗体内,加博沙多被迅速和完全吸收,可能在小肠近端,吸收后代谢最小。 联合给药色氨酸后,Gaboxadol/加博沙多的体内吸收[6] 同时给药hPAT1抑制剂色氨酸对加博沙多的吸收谱有剂量依赖性,导致Cmax降低和Tmax增加。加博沙多吸收率的降低可由胃排空率的改变引起。在人类中,胃排空的速度随着一餐所摄入的卡路里数量的增加而降低(Calbet和MacLean, 1997;Sunesen et al., 2005),在狗的实验中,膳食成分也被证明可以延长胃排空(Mizuta et al., 1990)。为了排除观察到的对加博沙多吸收的影响是胃排空改变的结果,研究了常被用作胃排空标志的扑热息痛的累积吸收曲线(Calbet and MacLean, 1997;Sunesen et al., 2005),在色氨酸存在的情况下进行了研究。高剂量色氨酸对扑热息痛的胃排空无显著影响。然而,色氨酸对加博沙多的ka有显著影响。由于共给色氨酸改变了加博沙多Tmax, Cmax和ka,而Fa, ke和AUC不变,色氨酸的作用可能是色氨酸和加博沙多在吸收部位相互作用的结果,而不是由于胃排空的改变。其他研究表明,加博沙多与血浆蛋白结合程度较低,不被细胞色素p -450代谢(Lund et al., 2006)。因此,根据体外实验结果表明,hPAT1介导了Caco-2单层中大部分加博沙多的腔内转运,似乎体内观察也可以解释为pat1介导的狗对加博沙多的吸收,而这种吸收被色氨酸的共同给药所减少。从色氨酸对加博沙多Cmax的影响或对肠道吸收速率常数ka的影响来估计色氨酸抑制加博沙多肠道运输的体内亲和值。IC50值分别为10.1和12.6 mmol·L−1。如前所述,在Caco-2细胞中,hPAT1对色氨酸的体外亲和力(通过hPAT1抑制脯氨酸运输)为7.7 mmol·L−1。考虑到色氨酸的抑制作用是针对两种不同的化合物(脯氨酸和加博沙多)测量的,并且体内转运不仅包括腔内转运,还包括体循环中的外观,其中遇到了加博沙多跨几个膜的转移,IC50值彼此惊人地接近。hPAT1底物的特点是在毫摩尔范围内具有亲和力,并且在整个肠道中表达的转运蛋白具有高容量(Chen et al., 2003)。加博沙多和色氨酸的摩尔给药比高达1:41,由于它们对hPAT1的亲和力相当,Cmax和ka的降低可能是由于加博沙多和色氨酸在吸收部位,即小肠肠细胞管腔膜上的PAT1蛋白处的竞争性相互作用。因此,最大血浆浓度随着Tmax的延长而出现,但由于容量过大和肠道中PAT1的表达,随着吸收沿着肠道的长度进行,吸收分数保持不变。因此,加博沙多的峰值血浆浓度可以通过修改吸收过程来降低,如这里所示,或者通过更经典的缓释制剂方法,如前面所建议的(Kjaer和Nielsen, 1983)。 |
| 细胞实验 |
细胞活力测定[1]
细胞类型:第 2/3 层神经元(来自雄性 C57BL/6 小鼠,出生后 13-19 天) 测试浓度: 1 µM 孵育时间: 5 s 实验结果: 诱导破伤风电流为 43.2 pA。 细胞活力测定[1] 细胞类型:第 2/3 层神经元(来自雄性 C57BL/6 小鼠,出生后 13-19 天) 测试浓度: 1 µM 孵育时间: 1 s 实验结果: 仅 mIPSC 的振幅和频率发生微小变化。 Caco-2细胞体外经上皮转运[2] 细胞培养和体外转运实验如前所述进行(Larsen等人,2008年)。简单地说,将第24 ~ 29代的细胞接种到Transwell™植入物中,并在播种后的第25 ~ 28天进行实验。在HBSS缓冲液中,测量了Gaboxadol/加博沙多在Caco-2细胞单层上从根尖到基底外侧(A-B)或基底外侧到根尖(B-A)方向的上皮转运。在所有实验中,基底外侧的HBSS的pH值为7.4,而顶部缓冲液在加入测试化合物后调整为pH 6.0或7.4。顶腔和底外侧腔体积分别为0.5和1.0 ml。在供体腔中加入新鲜的0.34、3.5或7.0 mM的加博沙多溶液开始运输实验。在18.8 mM 5-HTP存在和不存在的情况下,测定了加博沙多的转运。用18 μM [14C]-甘露醇测定了甘露醇在A-B方向的输运。分别于20、40、60、80和120 min后,从基底外侧和根尖接收室取100 μl和20 μl的样品。 经roat注射的卵母细胞摄取Gaboxadol/加博沙多[2] 在BD Gentest™运输细胞中研究了rOat1对加博沙多的摄取;预注射rOat1 cRNA的爪蟾卵母细胞。运输细胞按生产说明书处理。作为阴性对照,在注射水的卵母细胞中,即不表达rOat1的卵母细胞中,也测量了加博沙多的摄取。每个数据点5 - 10个卵母细胞放置在5ml试管中。卵母细胞用1 ml的Na+缓冲液洗涤3次,然后用100 μl的复合溶液孵育。在rOat1 cRNA注入的卵母细胞和对照卵母细胞中,分别于15、30、60、90 min和30、60、90 min取样,观察100 μM加博沙多摄取的时间依赖性。研究了10、50、100、300、500和1000 μM加博沙多在rOat1 cRNA注射的卵母细胞中60 min的摄取情况,同时研究了100、500和1000 μM加博沙多在水注射的卵母细胞中的摄取情况。测定rOat1 cRNA注射卵母细胞和水注射卵母细胞在5- htp、5-脯氨酸、1 mM PAH或1 mM probenecid存在下对100 μM加博沙多的摄取情况,孵育60 min后,卵母细胞在冰冷的Na+缓冲液中洗涤3次,每个卵母细胞置于Eppendorf管中,加入30 μl MilliQ水和25 μl内标。卵母细胞保存在- 80°C直到定量时间,通过解冻和旋转混合来裂解卵母细胞。在10,000 × g离心15分钟后,从上清中提取加博沙多,按照以下步骤进行分析:用250 μl冷(4°C)乙腈沉淀蛋白质,将干燥的样品在60 μl (30:70) MeOH/MeCN中重新溶解。 定量血浆中加博沙多/Gaboxadol和缓冲[2] 用液体萃取法从血浆和HBSS样品中提取加博沙多。将100 μl HBSS (80 μl纯净水加入20 μl样品中)或100 μl血浆样品与25 μl内标(d4-加博沙多)和25 μl纯净水混合。加入400 μl冷乙腈进行蛋白沉淀。10000 × g离心15 min后,将425 μl上清转移至玻璃管中,在45℃氮气下蒸发干燥。样品溶解于80 μl甲醇/乙腈(30:70)中,旋转混合10 min, 3000 rpm离心3 min。在盲法血浆中加入标准品,制备与血浆样品相似的标准品。采用亲水性相互作用色谱法对加博沙多进行定量,然后采用(Kall et al., 2007)改进的方案进行质谱/质谱检测。LC系统由安捷伦1100系列泵和脱气器组成。采用Phenomenex公司的Asahipak氨基柱(nh2p - 50,150 mm × 2mm),流动相为20.0 mm乙酸铵,pH值为4:乙腈(30:70),流速为0.2 ml/min。将20 μl样品进样于柱上,室温保存。总运行时间为10分钟,前5分钟的洗脱让浪费。加博沙多的洗脱时间约为8分钟。所使用的质谱联用系统包括Sciex API 4000质谱联用检测器,带有涡轮离子喷雾和涡轮V源(Applied Biosystems, CA, USA)。采用多反应监测(MRM)负电离方式检测加博沙多(前体139.1 Da,产物110.1 Da)和d4-加博沙多(前体143.0,产物112.2 Da)。在1.7 ~ 1000.0 ng/ml浓度范围内,信号呈线性关系,定量限为1.7 ng/ml。软件来自Analyst™。 细胞培养和体外实验方案如前所述(Larsen等人,2008年)。将第20 ~ 29代Caco-2细胞接种到Transwell™插入物(1.12 cm2, 0.4µm孔径)上,于接种后第25 ~ 28天进行实验。在汉克斯平衡盐溶液(HBSS)缓冲液中,测定了Gaboxadol/加博沙多在Caco-2细胞单分子层从根尖向基底侧方向(A-B)和基底侧向根尖方向(B-A)的顶端摄取和上皮转运。在所有实验中,基底外侧的缓冲液pH为7.4。除非另有说明,在加入Gaboxadol/加博沙多盐酸或35 mmol·L−1色氨酸后,将应用于顶室的缓冲液调至pH 6.0。研究了0.34、3.5或7.0 mmol·L−1Gaboxadol的转运情况。这些浓度的选择是基于人单次睡前口服15mg加博沙多,提供约0.34 mmol·L−1的通径浓度,以及所获得的加博沙多对hPAT1的亲和力。在根尖室中加入含有12.5 nmol·L−1(0.5µCi) L-(3H)脯氨酸和0-30 mmol·L−1加博沙多或0-35 mmol·L−1色氨酸的新鲜根尖HBSS培养基,进行根尖摄取实验。5 min后终止根尖摄取实验,对样品进行闪烁计数分析。[6] |
| 动物实验 |
动物/疾病模型: SD(Sprague-Dawley)大鼠(255-276 g)[2]。
剂量: 2.5 mg/kg(静脉注射);0.5 和 5.0 mg/kg(口服)。 给药途径: po(灌胃);静脉注射;单次给药。 实验结果: THIP 4 在 SD(Sprague-Dawley)大鼠中的药代动力学/PK 参数 1.19 [2]。 IV (2.5 mg/kg) PO (0.5 mg/kg) PO (5.0 mg/kg) AUC (ng/mL·h) 1193 263 1988 Tmax (h) - 0.22 0.33 Cmax (ng/mL) 4350 291 2061 CL/Fa (mL/h/kg) - 1939 2558 CL (mL/h/kg) 2043 - - Fa (%) 100 110 83 Sprague-Dawley 大鼠体内 Gaboxadol 的吸收 [2] 实验前,雄性 Sprague-Dawley 大鼠饲养并适应环境 7 天。实验当日,动物体重为 255–276 g。实验期间,大鼠饲喂标准饲料和饮水,直至给药前16-20小时停止进食。实验开始前及给药后2小时,大鼠均可自由饮水。除非另有说明,每只大鼠随机分配至静脉注射组或口服组,共设置6个平行组,每组6只大鼠(n = 6)。口服溶液中加博沙多浓度为0.5或5.0 mg/kg,静脉注射溶液中加博沙多浓度为2.5 mg/kg。大鼠口服给药剂量为10.0 ml/kg(即浓度为0.35和3.5 mM的加博沙多),静脉注射给药剂量为5 ml/kg(3.5 mM)。在口服等渗盐水或100.0 mg/kg 5-羟色氨酸(用于静脉注射加博沙多)或200.0 mg/kg 5-羟色氨酸(用于口服加博沙多)30分钟后,通过灌胃(10.0 ml/kg)给予加博沙多。所有溶液均在用Vapro蒸汽压渗透压计检测渗透压前调节pH至5.2,并用甘露醇调节至等渗。通过尾静脉穿刺采集0.2 ml血液样本,并收集到含有20 IU肝素的Eppendorf管中。分别在加博沙多给药后5、15、30、45、60分钟以及2、3、4、6、8小时采集血液样本。血浆立即通过3600 × g离心10分钟收集,并储存于−80 °C直至进一步分析。实验结束后,对动物实施安乐死。 大鼠胃排空研究[5] 采用与上述类似的方案,以对乙酰氨基酚为标记物,研究对乙酰氨基酚的吸收,并评估5-羟色氨酸(5-HTP)对大鼠胃排空率的影响。选取26只大鼠,随机分配接受5种不同配方的对乙酰氨基酚。大鼠分别接受120.0 mg/kg对乙酰氨基酚和2.5 mg/kg加博沙多(10 ml/kg)的静脉注射或口服溶液。口服溶液在预先口服等渗生理盐水、甘露醇或200 mg/kg 5-HTP 30分钟后给予。在口服等渗盐水(n = 6)或 200.0 mg/kg 5-HTP(n = 2)预给药 30 分钟后,静脉注射加博沙多。药物给药、血液采样和动物处理均按上述方法进行。动物/疾病模型: Fmr1 KO2 小鼠(Fmr1 启动子和第一个外显子缺失,导致小鼠 mRNA 和蛋白质缺失)[5] 剂量: 0.5、1、1.5、2、3、4 或 5 mg/kg 给药途径:腹腔注射 (ip) 实验结果: Fmr1 KO2 小鼠的活动过度症状恢复正常。 同一笼内的小鼠注射相同剂量的Gaboxadol或载体,突变体和对照组小鼠分开饲养。所有小鼠均群养于塑料笼(35 × 30 × 12 cm)中,每笼 5 只,并在实验前适应动物房至少一周。室温(21 ± 2°C)、相对湿度(55 ± 5%)、12 小时光照/黑暗循环(上午 7 点至晚上 7 点开灯)和空气交换(每小时 16 次)均由自动控制。所有小鼠均可自由摄取食物和水。所有测试均在光照期进行,由一位对基因型和药物处理不知情的实验人员完成。[2] Gaboxadol 处理和实验时间线:Fmr1 KO2 小鼠在每次测试日行为测试前 30 分钟注射载体(0.9% 无菌生理盐水)或 Gaboxadol(0.5、1、1.5、2、3、4 或 5 mg/kg,腹腔注射),每次测试间隔三天,以避免药物的累积效应。所有实验中均包含在同一时间点注射载体的野生型小鼠。对小鼠(每组 n = 10)进行行为学筛选,测试顺序如下,每次测试间隔 2-3 天:旷场测试(OFT;第 1 天)、连续通道测试(第 4 天)、明暗箱测试(第 7 天)、社交测试和攻击性测试(第 10 天)以及自我梳理和刻板行为测试(第 12 天)。[5] 犬对Gaboxadol的吸收[6] 所有动物护理和实验研究均已获得丹麦司法部指定的动物福利委员会的批准,并符合欧盟指令 86/609/EEC、丹麦动物实验法规以及美国国立卫生研究院《实验动物护理和使用指南》。选取六只成年雄性比格犬(体重15.9–21.7 kg),采用罗马象限设计随机分组,分别接受六种不同配方的加博沙多盐酸盐,试验持续六周。试验开始前,所有犬只禁食20–24小时,给药10小时后恢复喂食。加博沙多的给药途径有两种:静脉注射(1.0 mL·kg−1)或灌胃(5.0 mL·kg−1),后者通过软管直接灌胃给药。所有犬只均接受2.5 mg·kg−1的加博沙多。除加博沙多外,口服制剂中还分别含有0、2.5、10、50或150 mg·kg−1的色氨酸,以确保两种化合物同时发挥作用。所有溶液的pH值均调整至5.2,并使用Vapro蒸汽压渗透压计(型号552O,Wescor公司,美国犹他州洛根市)检测渗透压。静脉输液用葡萄糖调节至等渗。通过单次静脉穿刺从头静脉采集2 mL血样,并收集到含有200 IU肝素作为抗凝剂的Eppendorf管中。分别在给予加博沙多之前以及给药后5、15、30、60、90分钟、2、3、4、6、8和10小时采集血样。立即将血样在4–8°C下以2200 g离心15分钟,收集血浆,并储存于−80°C直至进一步分析。动物在两次治疗之间有6天的洗脱期。 犬胃排空的研究[6] 采用与之前描述的以对乙酰氨基酚为标记物的方案类似的方法,评估色氨酸对犬胃排空率的影响。选取6只犬(体重16.1–21.5 kg),随机分配接受三种不同配方的对乙酰氨基酚,进行交叉研究。犬分别接受50 mg·kg−1的对乙酰氨基酚,以静脉注射(1 mL·kg−1)或口服溶液(5 mL·kg−1)的形式给药,该溶液含有2.5 mg·kg−1的Gaboxadol和0或150 mg·kg−1的色氨酸。犬只禁食、给药、采血和洗脱均按先前所述进行。 分析方法[6] 血浆和缓冲液中Gaboxadol的定量:采用液液萃取法从血浆和缓冲液样品中提取Gaboxadol。将100 µL HBSS或血浆样品与25 µL内标物(d4-gaboxadol)和25 µL纯水混合。加入400 µL冷乙腈进行蛋白质沉淀。10000 g离心15分钟后,取425 µL上清液转移至玻璃管中,并在45℃氮气流下蒸干。将样品重新溶解于80 µL甲醇/乙腈(30:70)混合液中,涡旋混合10分钟,并在3300×g下离心3分钟。随后采用亲水相互作用色谱-串联质谱(MS/MS)法对加博沙多进行定量分析,该方法参考了Kall等人(2007)的方法并进行了改进。液相色谱(LC)系统由安捷伦1100系列泵和脱气机组成。色谱柱为Phenomenex公司(美国加利福尼亚州托兰斯市)的Asahipak氨基柱(NH2P-50,150 × 2 mm),流动相为20.0 mmol·L⁻¹乙酸铵(pH 4)-乙腈(30:70),流速为0.2 mL·min⁻¹。进样量为20 μL,色谱柱保持在室温下。总运行时间为10 min,前5 min的洗脱液弃去。加博沙多在色谱柱上的洗脱时间约为8 min。所用的MS/MS系统由一台配备Turbo Ion Spray和Turbo V离子源的Sciex API 4000 MS/MS检测器(Applied Biosystems,美国加利福尼亚州福斯特城)组成。信号在0.5至2500 ng·mL⁻¹范围内呈线性关系,该方法的定量限为0.5 ng·mL⁻¹。分析软件为Analyst™(Applied Biosystems,版本4.0)。 |
| 药代性质 (ADME/PK) |
Sprague-Dawley大鼠口服Gaboxadol的药代动力学分析[2]
图1显示了Sprague-Dawley大鼠静脉注射和口服Gaboxadol后的血浆浓度曲线。0.5 mg/kg和5.0 mg/kg剂量组的血浆峰浓度时间(Tmax)分别为0.2 h和0.3 h(表2)。0.5 mg/kg剂量组的口服吸收似乎完全,平均生物利用度(Fa)为110%。5 mg/kg剂量组的Fa为83 ± 5%,显著低于0.5 mg/kg剂量组的Fa(p < 0.01)。加博沙多的清除率(CL/Fa)在0.5 mg/kg和5 mg/kg剂量下分别从1.93 L/h/kg增加到2.56 L/h/kg(p < 0.05)。当剂量从0.5 mg/kg增加到5 mg/kg(增加10倍)时,血浆峰浓度(Cmax)增加了7倍,表明在所研究的剂量范围内,峰浓度与剂量不呈比例关系。此外,剂量增加10倍后,AUC仅增加了7.5倍。非线性药代动力学表明,加博沙多的吸收或消除过程可能涉及载体介导的转运。该载体可能是肠道中的饱和吸收载体,也可能是参与肾脏对加博沙多重吸收的载体。对Sprague-Dawley大鼠口服加博沙多和5-羟色氨酸[2] 静脉和口服给药后,5-羟色氨酸的给药显著改变了大鼠血浆中加博沙多的浓度曲线(图1A-C)。吸收和消除相均发生改变,此外,预先给予5-羟色氨酸显著增加了大鼠的AUC。在加博沙多的初始吸收阶段(≤Tmax),与未接受5-羟色氨酸给药的动物相比,接受5-羟色氨酸给药的动物血浆中加博沙多的浓度显著降低(p < 0.05,图1B和C)。 5-HTP 对加博沙多尔的血浆峰浓度(Cmax)无显著影响。此外,5-HTP 显著延长了 0.5 mg/kg 和 5.0 mg/kg 剂量下加博沙多尔的达峰时间(Tmax)(p < 0.01 和 p < 0.001),表明 5-HTP 可改变大鼠肠道对加博沙多尔的吸收。图 1A 和 1B 的半对数图中显示,给予 5-HTP 的大鼠加博沙多尔消除曲线近似呈直线,而单独给予加博沙多尔的大鼠则呈双相曲线。5-HTP 的存在使 0.5 mg/kg 和 5.0 mg/kg 剂量下加博沙多尔的平均 AUC 分别增加了 5.5 倍(p < 0.01)和 3.6 倍。 0.5 mg/kg 和 5.0 mg/kg 加博沙多的平均清除率 CL/Fa 分别降低了 77% (p < 0.01) 和 23% (无统计学意义)。在 5-HTP 存在的情况下,2.5 mg/kg (IV) 加博沙多的清除率 CL 降低了 66% (n = 2)(表 2)。本研究中,对乙酰氨基酚的口服吸收(作为胃排空的标志物)在预先给予 5-HTP 或甘露醇后,根据 Cmax、Tmax、AUC、Fa 和 CL/Fa 的评估结果,未发生显著变化(图 2 和表 3)。因此,可以得出结论,5-HTP 不会显著延长胃排空。然而,对乙酰氨基酚的消除速率常数 ke (0.95 h−1) 在 5-HTP 存在下显著降低 (0.53 h−1, p < 0.05)。加博沙多是 rOat1 的底物,但 5-HTP 不是 [2] 由于 5-HTP 对大鼠体内加博沙多的清除率有显著影响,因此在有或无 5-HTP 的情况下,研究了肾脏有机阴离子转运蛋白 rOat1 对加博沙多的转运(图 3,图 4)。在所研究的 90 分钟内,rOat1 cRNA 注射组和注射水组的卵母细胞对加博沙多的摄取均呈线性关系(图 3A)。rOat1 cRNA 注射组卵母细胞对加博沙多的摄取量显著高于注射水组卵母细胞(p ≤ 0.001)。在注射了rOat1 cRNA的卵母细胞中,gaboxadol的浓度依赖性摄取速率呈饱和性(图3B)。数据点符合米氏动力学,Km值为151.2 ± 58.19 μM,Vmax值为0.78 ± 0.09 pmol oocyte−1 min−1。因此,gaboxadol是rOat1的底物。我们随后研究了在5 mM 5-HTP、5 mM L-脯氨酸、1 mM PAH和1 mM丙磺舒存在下,rOat1介导的gaboxadol摄取情况(图4)。 5-HTP 或 L-脯氨酸的存在并不影响加博沙多尔的摄取,而 PAH 和丙磺舒(分别为 Oat1 的底物和抑制剂)的存在则显著降低了加博沙多尔的摄取(p < 0.001)。5-HTP 在体外和体内均能降低加博沙多尔的吸收[2]。加博沙多尔在 Caco-2 细胞单层上的转运呈 A-B 方向极化。A-B 方向的加博沙多尔转运取决于供体浓度和供体培养基的 pH 值。这与加博沙多尔通过 hPAT1 介导的跨肠细胞腔膜转运相一致,该转运步骤对于最终的跨上皮吸收至关重要。在Caco-2细胞单层中也观察到了类似的现象,即其他PAT1底物(如L-脯氨酸(Thwaites等,1993)和GABA(Thwaites等,2000))的pH依赖性转运。此前,Chen等从Caco-2细胞中克隆了hPAT1,免疫荧光显示PAT1表达于Caco-2细胞和大鼠小肠肠上皮细胞的顶膜(Chen等,2003;Anderson等,2004)。在Caco-2细胞单层中,80-90%的加博沙多转运很可能是由hPAT1介导的,而5-HTP抑制了这种转运。加博沙多和5-羟色氨酸通过hPAT1抑制Caco-2细胞摄取L-脯氨酸的Ki值分别为6.6 mM(Larsen等人,2009)和2.3 mM(Larsen等人,2008)。因此,问题在于在相关浓度下是否能观察到加博沙多和5-羟色氨酸之间的相互作用。假设口服加博沙多剂量为15 mg,体积为250 ml,则管腔内浓度为0.34 mM。5-羟色氨酸的常用剂量为50-100 mg,管腔内浓度为9-18 mM。在本实验中,加博沙多的最低浓度为0.34 mM,5-羟色氨酸的最低浓度为18.8 mM。因此,可以在体内测定这些浓度,并且两种化合物之间的相互作用可能具有相关性。在体内,口服给予Sprague-Dawley大鼠后,血浆中加博沙多尔的峰值浓度在0.22-0.33小时内达到。在加博沙多尔的初始吸收阶段,与仅服用加博沙多尔的大鼠相比,预先服用5-HTP的大鼠血浆中加博沙多尔的浓度显著降低。此外,5-HTP的存在显著延长了加博沙多尔的达峰时间(Tmax)。一项平行研究对乙酰氨基酚的吸收(乙酰氨基酚是常用的胃排空标志物)表明,5-HTP并未显著延缓胃排空。这表明5-HTP降低了大鼠小肠中加博沙多尔的吸收速率。结合体外hPAT1介导的加博沙多跨上皮转运的研究结果,提示5-HTP可能通过与大鼠小肠中的PAT1相互作用降低加博沙多的吸收率。4.2. 加博沙多的肾脏排泄可能依赖于转运蛋白和5-HTP的存在。除了降低加博沙多的初始吸收外,5-HTP还增加了AUC,这主要是由于其改变了加博沙多在达峰时间(Tmax)后的血浆浓度曲线。既往研究表明,加博沙多的血浆蛋白结合率较低(Lund等,2006),并且它不是细胞色素P-450的底物(Schultz等,1981;Lund等,2006)。因此,加博沙多清除率的降低可能是导致其AUC增加的原因。Tmax显著延长与加博沙多清除率降低相符;然而,5-HTP预给药并未显著改变Cmax。或许,rPat1介导的加博沙多吸收率抑制和清除率降低同时作用,可以解释观察到的稳定的Cmax水平。加博沙多主要经肾脏排泄,尿液中可检测到不同量的代谢物。在人尿中,34%的加博沙多以O-葡萄糖醛酸苷的形式存在(Lund等,2006)。然而,在大鼠尿液中仅检测到2-7%的O-葡萄糖醛酸苷,因此代谢变化似乎并不影响本研究的结果(Schultz等人,1981)。 在确定rPat1是最有可能负责Gaboxadol肠道转运的转运蛋白后,我们寻找了可能参与肾脏对gaboxadol处理的其他转运蛋白。质子偶联氨基酸转运蛋白rPat1和rPat2可能参与尿液中gaboxadol的重吸收,但5-HTP的作用可能是降低重吸收,从而增加肾脏排泄和清除。预先给予5-HTP后,观察到相反的结果,即CL和CL/Fa均降低,这表明大鼠肾上皮细胞顶膜上的外排转运蛋白或基底外侧膜上的内流转运蛋白可能受到抑制。先前一项关于阿昔洛韦在人体内的药代动力学和消除的研究发现,当患者同时服用丙磺舒时,会出现类似的AUC增加和药物排泄率降低的现象(Laskin等,1982)。由于丙磺舒是OAT1等外排转运蛋白的抑制剂,而阿昔洛韦是rOAT1的底物,因此二者在hOAT1上的相互作用很可能导致药物的肾小管分泌减少(Wada等,2000)。此外,最近有研究表明,加博沙多进入人肾脏的转运是通过hOAT1进行的(Chu等,2009)。因此,我们研究了大鼠肾脏基底外侧有机阴离子转运蛋白1 (rOat1),结果表明加博沙多是其底物,而5-羟色氨酸(5-HTP)则不是。有机阴离子转运蛋白具有广泛的底物特异性,已被证明能够将尤其是小分子两亲性物质转运穿过边界上皮细胞(Rizwan和Burckhardt,2007)。在注射了rOat1 cRNA的卵母细胞中,加博沙多的摄取速率以平均Km值为151 μM为特征。该值与在转染了hOAT1的CHO-K1细胞中获得的加博沙多摄取Km值(115 ± 27 μM)非常接近(Chu等,2009)。相比之下,大多数抗生素对hOAT1的亲和力较低,而非甾体抗炎药(NSAIDs)的亲和力较高,因此加博沙多尔被认为是rOat1的中等亲和力底物(Apiwattanakul等,1999;Rizwan和Burckhardt,2007)。rOat1的鼠源同源物mOat1和hOAT1已被观察到能够促进神经活性色氨酸代谢物的转运(Alebouyeh等,2003;Bahn等,2005)。然而,5-羟色氨酸(5-HTP)的存在并未降低卵母细胞通过rOat1对加博沙多尔的摄取,表明5-HTP并不直接抑制加博沙多尔通过rOat1的排泄。然而,在大鼠体内,5-HTP 可被芳香族 L-氨基酸脱羧酶 (LAAD) 代谢为 5-羟色胺(血清素或 5-HT),并进一步被单胺氧化酶 (MAO) 代谢为 5-羟基吲哚乙酸 (5-HIAA) (Stier 等,1984;Wang 等,2001)。据报道,1 mM 5-HIAA 可抑制 mOat1 转染的 COS-7 细胞对 0.25 μM [3H]-PAH 的摄取约 90% (Bahn 等,2005)。因此,根据大鼠体内5-HIAA的生成速率和化合物的亲和力,5-HIAA可能与rOat1上的加博沙多发生相互作用。 在本体内研究中,给予5.0 mg/kg剂量后观察到的加博沙多血浆最大浓度约为2000 ng/ml,相当于14.3 μM。因此,加博沙多的清除可能不受rOat1转运能力的限制。5-HTP对加博沙多清除率的影响无法用本文鉴定的转运蛋白来解释。一种可能的解释是5-HT对肾血流量和肾小球滤过率的影响。在离体灌注大鼠肾脏以及活体大鼠肾脏中,5-羟色胺(5-HT)已被证实可介导入球小动脉的选择性收缩,并显著改变肾功能(Stier等,1984;Ding等,1989;Wang等,2001)。在麻醉大鼠中,以15和75 μg/min的速率持续输注5-羟色氨酸(5-HTP)20分钟后,肾小球滤过率、有效血浆流量和尿流量分别下降了26%~41%(Stier和Itskovitz,1985)。这也可以解释本研究中观察到的对乙酰氨基酚消除速率常数降低的现象,并且通常也适用于具有一定程度肾脏排泄的化合物,尽管不能排除Oat1受体的相互作用(Khamdang等,2002)。 [2] 结论 本研究表明,Gaboxadol在Caco-2细胞单层上的转运很可能是通过PAT1介导的,而5-HTP会抑制这种转运。大鼠口服Gaboxadol的吸收也可能由rPat1介导,因为5-HTP的存在会降低Gaboxadol的初始吸收。阴离子转运蛋白rOat1被认为参与了大鼠肾脏对Gaboxadol的处理。本研究揭示了一种可能的药物-膳食补充剂相互作用,因为5-HTP显著改变了Gaboxadol的整体药代动力学特征。虽然很难推测人类服用Gaboxadol治疗失眠或其他疾病时同时服用5-HTP补充剂会产生什么后果,但它可能会导致严重的副作用。鉴定参与加博沙多药代动力学特征的转运蛋白,为进一步了解潜在的药物相互作用奠定了基础。 犬口服加博沙多吸收的药代动力学分析[6] 在比格犬中,口服或静脉注射2.5 mg kg−1加博沙多后,监测了10小时内的血浆浓度曲线(图3)。犬口服加博沙多后的生物利用度Fa较高(超过80%)(表1)。口服联合使用2.5–150 mg kg−1色氨酸并未显著改变加博沙多的AUC,且各制剂的平均相对生物利用度在75%(10 mg kg−1色氨酸)至86.1%(2.5 mg kg−1色氨酸)之间。此外,加博沙多尔的消除速率常数 (ke) 和清除率 (CL) 并未因与色氨酸合用而改变。然而,与色氨酸合用 150 mg kg⁻¹ 后,加博沙多尔的血浆峰浓度 (Cmax) 从 2502 ng mL⁻¹ 降至 1419 ng mL⁻¹,即降低了 57%。此外,达到血浆峰浓度所需的时间 (Tmax) 从 0.46 h 延长至 1.5 h (P < 0.01)。随后,将五个剂量组的 Cmax 值拟合到剂量-反应曲线(图 4),结果表明加博沙多尔的吸收与色氨酸浓度之间存在直接相互作用。色氨酸对加博沙多Cmax的体内IC50值估计为12.6 mg kg−1,相当于12.3 mmol L−1的色氨酸浓度(未校正胃肠液稀释)。 加博沙多和对乙酰氨基酚的吸收速率常数[6] 如图5A所示的反卷积曲线所示,同时给予递增剂量的色氨酸逐渐改变了加博沙多的平均累积吸收分数。与单独服用加博沙多相比,在150 mg kg−1色氨酸存在下,加博沙多在0.5-1.25小时时间点的吸收显著降低。口服对乙酰氨基酚60分钟后吸收率为91.5 ± 3.3%(图5B),表明胃排空主要发生在给药后的第一个小时内。同时给予 150.0 mg kg−1 色氨酸并未显著改变胃排空率,因为在测试的时间点,无论是否存在色氨酸,对乙酰氨基酚的吸收分数均无显著差异。血浆中对乙酰氨基酚浓度的药代动力学参数Tmax、AUC和CL与对乙酰氨基酚和色氨酸联合给药后获得的参数无显著差异(结果未显示)。基于图5A所示的曲线,计算了加博沙多尔的吸收速率常数ka,并将其作为色氨酸剂量对数的函数绘制在图6A中。与色氨酸联合给药可降低加博沙多尔的ka值,其体内IC50值为10.3 mg kg−1,相当于浓度为10.1 mmol L−1的色氨酸口服溶液。图6B显示,150 mg kg−1的PAT1抑制剂色氨酸显著降低了加博沙多尔的吸收速率常数(P < 0.01),而对乙酰氨基酚的吸收速率常数无显著影响。 |
| 毒性/毒理 (Toxicokinetics/TK) |
小鼠腹腔注射 LD50 98 mg/kg 感觉器官和特殊感觉:眼睑下垂;行为:嗜睡(总体活动抑制);皮肤及其附属器官(皮肤);毛发;其他 神经药理学,21(803),1982 [PMID:7121752]
|
| 参考文献 |
|
| 其他信息 |
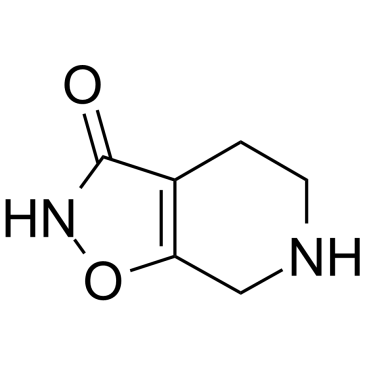
4,5,6,7-四氢异噁唑并[5,4-c]吡啶-3-醇是一种噁唑类化合物。
加博沙多(Gaboxadol),又名4,5,6,7-四氢异噁唑并[5,4-c]吡啶-3-醇(THIP),是由灵北制药和默克公司开发的一种实验性助眠药物。据报道,该药物可以增加深度睡眠,且不具有苯二氮卓类药物的强化作用。由于对安全性和有效性的担忧,加博沙多的研发于2007年3月停止。它作用于GABA系统,但其作用机制似乎与苯二氮卓类药物和其他镇静剂不同。 药物适应症 已研究用于治疗睡眠障碍和失眠。 我们研究了δ亚基优先性催眠药物THIP对小鼠新皮层神经元GABAA受体的作用。我们发现,在临床相关浓度下,Gaboxadol/THIP 可诱导新皮层神经元产生显著的 GABAA 受体介导的强直电流,而不影响突触内 GABAA 受体。该强直电流的强度与新皮层各层中 δ 亚基的表达呈定性相关。出乎意料的是,THIP 显著降低了第 2/3 层神经元中由活动驱动的自发性抑制性突触后电流 (sIPSC) 的频率,同时增加了自发性兴奋性突触后电流 (sEPSC) 的频率。这些结果表明,THIP 主要作用于新皮层的突触外 GABAA 受体,并且可能通过抑制中间神经元活动来减少抑制性突触输入。这或许可以解释为何THIP会改变皮层振荡活动(Vyazovskiy等人,2005),因为神经元放电时间部分取决于广泛连接的中间神经元网络(Tamas等人,2000;Buhl等人,1998)。[1] GABA(C)受体在神经系统功能的诸多方面(包括记忆、近视、疼痛和睡眠)中的作用正受到广泛研究。已有证据表明,视网膜、海马、脊髓、上丘、垂体和肠道等多种组织中存在功能性GABA(C)受体。本文综述了多种神经化学物质,这些物质已被证明可用于区分GABA(C)受体与其他主要抑制性神经递质GABA的受体。本文描述了一些选择性激动剂(包括(+)-CAMP和5-甲基-IAA)、竞争性拮抗剂(例如TPMPA、(±)-顺式-3-ACPBPA和aza-THIP)、正向调节剂(别孕烷醇酮)和负向调节剂(表孕烷醇酮、洛瑞克唑)。此外,还介绍了可能有助于区分同源ρ1和ρ2 GABA(C)受体的神经化学物质(2-甲基-TACA和环噻嗪)。由于GABA(C)受体与GABA(A)和GABA(B)受体相比分布范围较小、丰度较低且结构相对简单,因此它们是极具吸引力的药物靶点。[4] Gaboxadol在0.5 mg/kg剂量下可使Fmr1 KO2小鼠所有测试的行为缺陷恢复正常。虽然更高剂量也能使易激惹和攻击性行为恢复正常,但在其他评估的行为领域中未观察到这种现象。此处观察到的疗效窗口略窄,一种解释可能源于先前的研究表明,信息处理受损是由于持续性抑制不足或过度所致,而Gaboxadol正是增强这种生理过程。根据这一模型,高剂量药物带来的行为益处会被药物引入的、与脆性X综合征无关的缺陷所抵消(Duguid等人,2012)。我们的研究结果为Gaboxadol在逆转自闭症谱系障碍相关行为、攻击性和社交能力方面的潜在益处提供了强有力的证据。综上所述,这些结果支持以下假设:Gaboxadol增强突触外GABAA受体的活性可能对脆性X综合征患者有益。总之,这些数据支持未来对脆性X综合征(FXS)患者进行加博沙多(gaboxadol)的评估,尤其是在多动、焦虑、自闭症谱系障碍(ASD)相关刻板行为、社交能力、易怒、攻击性和认知功能等方面的症状。[5] 背景和目的:加博沙多(gaboxadol)一直处于研发阶段,用于治疗慢性疼痛和失眠。临床应用表明,加博沙多(gaboxadol)的不良反应似乎与血清峰浓度相关。本研究旨在体外和体内研究加博沙多(gaboxadol)的肠道吸收机制。 实验方法:体外转运研究在Caco-2细胞单层中进行。体内药代动力学研究在比格犬中进行。给予受试者2.5 mg·kg⁻¹的Gaboxadol,分别采用静脉注射(1.0 mL·kg⁻¹)或口服溶液(5.0 mL·kg⁻¹)的方式给药。主要结果:Gaboxadol可能是人质子偶联氨基酸转运蛋白hPAT1的底物,它能抑制Caco-2细胞单层中hPAT1介导的L-[³H]脯氨酸的摄取,其抑制常数K₁为6.6 mmol·L⁻¹。Gaboxadol的跨上皮转运呈顶端至基底外侧方向的极化,且依赖于Gaboxadol的浓度和顶端缓冲溶液的pH值。在比格犬中,加博沙多的吸收几乎完全(绝对生物利用度 F(a) 为 85.3%),达峰时间 (T(max)) 为 0.46 小时。口服联合使用 2.5-150 mg·kg⁻¹ 的 PAT1 抑制剂 L-色氨酸,可显著降低加博沙多的吸收速率常数 k(a) 和 C(max),并延长其 T(max),而曲线下面积和清除率保持不变。 结论和意义:加博沙多通过小肠肠细胞腔膜的吸收可能由 PAT1 介导。这项研究有助于降低加博沙多尔的吸收率,从而降低血浆峰浓度。[6] 总之,本研究首次表明,加博沙多尔在Caco-2细胞单层上的高渗透性很可能是由于PAT1介导的跨腔膜转运,从而导致高跨上皮转运。体外加博沙多尔转运动力学和在犬体内观察到的药代动力学均支持以下结论:PAT1介导加博沙多尔在体外和体内均能跨黏膜转运。此外,本研究表明,可以利用转运蛋白的活性来调节或控制药物的肠道吸收。该制剂设计为降低加博沙多尔的血浆峰浓度提供了一种简便的方法,同时保持了较高的生物利用度。这可能有助于减少与高血浆峰浓度相关的副作用。[6] |
| 分子式 |
C₆H₈N₂O₂
|
|---|---|
| 分子量 |
140.14
|
| 精确质量 |
140.058
|
| CAS号 |
64603-91-4
|
| 相关CAS号 |
Gaboxadol hydrochloride;85118-33-8
|
| PubChem CID |
3448
|
| 外观&性状 |
White to yellow solid powder
|
| 密度 |
1.3±0.1 g/cm3
|
| 沸点 |
340.5±42.0 °C at 760 mmHg
|
| 闪点 |
159.7±27.9 °C
|
| 蒸汽压 |
0.0±0.8 mmHg at 25°C
|
| 折射率 |
1.551
|
| LogP |
-0.61
|
| tPSA |
58.29
|
| 氢键供体(HBD)数目 |
2
|
| 氢键受体(HBA)数目 |
3
|
| 可旋转键数目(RBC) |
0
|
| 重原子数目 |
10
|
| 分子复杂度/Complexity |
210
|
| 定义原子立体中心数目 |
0
|
| SMILES |
C1CNCC2=C1C(=O)NO2
|
| InChi Key |
ZXRVKCBLGJOCEE-UHFFFAOYSA-N
|
| InChi Code |
InChI=1S/C6H8N2O2/c9-6-4-1-2-7-3-5(4)10-8-6/h7H,1-3H2,(H,8,9)
|
| 化学名 |
4,5,6,7-Tetrahydroisoxazolo(5,4-c)pyridin-3(2H)-one4,5,6,7-Tetrahydro-[1,2]oxazolo[5,4-c]pyridin-6-ium-3-one
|
| 别名 |
Gaboxadol; OV-101; MK0928; Lu02030; OV101; Lu-02-030; gaboxadol; 64603-91-4; Gaboxadolum; Gaboxadolum [Latin]; Gaboxadol [USAN:INN]; 4,5,6,7-Tetrahydroisoxazolo(5,4-c)pyridin-3-ol; Lu 02-030; MK-0928; Lu-02030
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO : ~50 mg/mL (~356.79 mM)
H2O : ~23.33 mg/mL (~166.48 mM) |
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.5 mg/mL (17.84 mM) (饱和度未知) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 25.0 mg/mL澄清DMSO储备液加入到400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: ≥ 2.5 mg/mL (17.84 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 例如,若需制备1 mL的工作液,可将 100 μL 25.0 mg/mL 澄清 DMSO 储备液加入到 900 μL 玉米油中并混合均匀。 View More
配方 3 中的溶解度: ≥ 2.08 mg/mL (14.84 mM) (饱和度未知) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 配方 4 中的溶解度: 50 mg/mL (356.79 mM) in PBS (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液; 超声助溶. 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 7.1357 mL | 35.6786 mL | 71.3572 mL | |
| 5 mM | 1.4271 mL | 7.1357 mL | 14.2714 mL | |
| 10 mM | 0.7136 mL | 3.5679 mL | 7.1357 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
| NCT Number | Recruitment | interventions | Conditions | Sponsor/Collaborators | Start Date | Phases |
| NCT00209963 | Completed | Drug: Gaboxadol | Primary Insomnia | H. Lundbeck A/S | 2003-06 | Phase 3 |
| NCT06334419 | Recruiting | Drug: Gaboxadol Drug: Placebo |
Fragile X Syndrome | Craig Erickson | 2024-01-29 | Phase 2 |
| NCT00209846 | Completed | Drug: Gaboxadol | Primary Insomnia | H. Lundbeck A/S | 2004-06 | Phase 3 |
| NCT00209924 | Completed | Drug: Gaboxadol | Primary Insomnia | H. Lundbeck A/S | 2004-04 | Phase 3 |
| NCT02996305 | Completed | Drug: OV101 Regimen 1 Drug: OV101 regimen 2 Other: Placebo |
Angelman Syndrome | Ovid Therapeutics Inc. | 2016-01 | Phase 2 |
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1
































 463611831
463611831
