| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 1mg |
|
||
| 5mg |
|
||
| 10mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| Other Sizes |
|

| 靶点 |
Inhibits the interaction between eIF4E (cap-binding protein) and eIF4G (large scaffolding protein), as well as the interaction between eIF4E and 4E-BP1. In a TR-FRET assay monitoring eIF4E-eIF4GI interaction, IC50 = 13.5 μM [1]
Does not compete with 5′ mRNA cap structures for binding to eIF4E [1] |
|---|---|
| 体外研究 (In Vitro) |
4E2RCat 通过干扰主要支架蛋白 eIF4G 与帽结合蛋白 eIF4E 之间的相互作用来抑制帽依赖性翻译。它显著降低了由人冠状病毒 229E (HCoV-229E) 引起的感染细胞数量以及细胞内和细胞外感染性病毒滴度。4E2RCat 以剂量依赖的方式抑制帽依赖性翻译。4E2RCat 抑制帽依赖性 FF 翻译,而对 EMCV IRES 驱动的 Ren 翻译不受影响。4E2RCat 以时间和剂量依赖的方式抑制冠状病毒复制 [1]。
在用 FF/HCV/Ren 双顺反子 mRNA 编程的 Krebs 提取物中,4E2RCat 以剂量依赖的方式抑制帽依赖性翻译;在 25 μM 时,抑制 FF(帽依赖性)翻译,而对 HCV IRES 驱动的 Ren 翻译不受影响(甚至由于核糖体可用性而增加)。在 100 μM 浓度下,观察到对 HCV IRES 的一些脱靶效应 [1] - 在含有 FF/EMCV/Ren 双顺反子 mRNA 的同一系统中,4E2RCat 抑制依赖于帽结构的 FF 翻译,但不抑制 EMCV IRES 驱动的 Ren 翻译 [1] - 在使用重组 eIF4E 和 GST-eIF4GI517-606 的 pull-down 实验中,100 μM 的 4E2RCat 阻断了它们的相互作用 [1] - 100 μM 的 4E2RCat 也阻断了 eIF4E 与 GST-eIF4GII555-658 以及 eIF4E 与 GST-4E-BP1 之间的相互作用 [1] - 在使用核糖体盐洗 (RSW) 的 m7GTP pull-down 实验中,25 μM 的 4E2RCat 破坏了预先形成的 eIF4F 复合物:eIF4GI 和 eIF4A 与 eIF4E 的共纯化显著减少,而eIF4E 恢复保持不变 [1] - 通过 [35S]甲硫氨酸代谢标记测定,以剂量和时间依赖的方式抑制 L132 细胞中的蛋白质合成:处理 4 小时后,相对蛋白质合成在 6.25 μM 时约为 80%,在 12.5 μM 时约为 60%,在 25 μM 时约为 50%,在 50 μM 时约为 40%;处理 24 小时后,相对合成在 6.25 μM 时约为 60%,在 12.5 μM 时约为 40%,在 25 μM 时约为 30%,在 50 μM 时约为 15% [1] - 在 HCoV-229E 感染的 L132 细胞(MOI 0.1)中,12.5 μM 的 4E2RCat 在感染后 24 小时和 48 小时完全阻断细胞外感染性病毒的产生(低于检测限);在 6.25 μM 浓度下,48 小时后细胞外病毒浓度非常低 (<10 TCID50/ml) [1] - 细胞内感染性病毒滴度也显著降低:在 12.5 μM 4E2RCat 存在下,感染后 24 小时和 48 小时,滴度分别为 <10 TCID50/ml 和 <20 TCID50/ml,与 DMSO 对照组相比,降低了近 10^6 倍 [1] - 以剂量依赖的方式降低 HCoV-229E S 蛋白阳性细胞的百分比:在 6.25 μM 浓度下,降低约 2.4 倍;在 12.5 μM 浓度下,与对照组相比,细胞内蛋白质合成减少约 4 倍 [1] - 在 50 μM 浓度下,尽管 4E2RCat 能显著降低宿主细胞的蛋白质合成,但对感染 2 PFU/细胞的 1 型脊髓灰质炎病毒(Mahoney 株)的 HeLa 细胞中的脊髓灰质炎病毒蛋白合成没有影响 [1] - 对 19 种类似物进行结构-活性关系 (SAR) 分析,发现其中 4 种化合物具有一定的活性,但没有一种化合物在阻止 eIF4E-eIF4G 相互作用或阻断细胞蛋白质合成方面像 4E2RCat 那样有效 [1] |
| 体内研究 (In Vivo) |
4E2RCat 对体内蛋白质合成的抑制并非源于细胞死亡的增加[1]。
|
| 酶活实验 |
利用时间分辨荧光共振能量转移 (TR-FRET) 技术,采用微型化的 1536 孔板形式,对 eIF4E-eIF4G 抑制剂进行超高通量筛选。该技术可监测 eIF4E 和 eIF4GI 之间的相互作用。筛选了包含 217,341 个化合物的化合物库,并鉴定出 4E2RCat。在该 TR-FRET 检测中,4E2RCat 的 8 点剂量反应曲线显示 IC50 为 13.5 μM [1]。
- 为评估 eIF4E 结合伴侣的抑制情况,进行下拉实验:将 GST-eIF4GI517-606 或牛血清白蛋白 (BSA) 偶联到 Affi-Gel-10 磁珠上。将重组 eIF4E 与 100 μM 4E2RCat 或 DMSO 在室温下预孵育 1 小时,然后与 Affi-Gel 结合的蛋白质混合,再孵育 1 小时。洗涤磁珠,通过 SDS-PAGE 电泳分离,并通过 Western 印迹法检测 eIF4E。类似地,使用 2.5 μg GST 融合蛋白和 0.25 μg eIF4E 进行 GST-eIF4GII555-658 和 GST-4E-BP1 的下拉实验,在结合缓冲液中与 25 μM 4E2RCat 孵育,然后用谷胱甘肽珠洗脱,并通过 Western blot 分析 [1] - 为了评估对预先形成的 eIF4F 复合物的影响:将核糖体盐洗 (RSW) 与 25 μM 4E2RCat 或 DMSO 在 30°C 下孵育 1 小时,然后加入 50 μl 50% m7GTP Sepharose 珠,并在 4°C 下旋转 2 小时。将磁珠洗涤后,用 m7GTP 洗脱蛋白质,并使用抗 eIF4E、抗 eIF4GI 和抗 eIF4A 抗体通过 SDS-PAGE 和 Western blot 分析洗脱液 [1] - 对 eIF4E 三维结构进行计算机模拟溶剂映射,确定了五个浅口袋作为潜在的结合热点。使用 AutoDock Vina 1.1.0 对接 4E2RCat 的结果表明,4E2RCat 结合到五个口袋中的四个,预计会与 eIF4E-eIF4G 和 eIF4E-4E-BP1 相互作用发生冲突 [1] |
| 细胞实验 |
体内代谢标记:在处理前24小时,将L132细胞以60,000个细胞/孔的密度接种于24孔板中。细胞用递增浓度的4E2RCat处理4小时或24小时。在最后1小时,将培养基更换为不含甲硫氨酸的DMEM培养基,并添加10%透析血清;在最后15分钟,用[35S]甲硫氨酸(150-225 μCi/ml)标记细胞。洗涤细胞后,用RIPA裂解缓冲液裂解细胞,用三氯乙酸沉淀样品,并通过闪烁计数法测定放射性。测定蛋白质浓度并用于标准化计数[1]
- 细胞活力/凋亡检测:将L132细胞(6孔板中每孔200,000个细胞)用12.5 μM 4E2RCat处理指定时间(4、8、24、48、72小时)。收集细胞(包括上清液),洗涤后重悬于Annexin V结合缓冲液中,然后用Annexin V/碘化丙啶染色,并通过流式细胞术进行分析。相对于DMSO溶剂对照组(设定为1),凋亡细胞的比例在任何时间点均未见增加[1] - 冠状病毒感染和滴度测定:将L132细胞以0.1的MOI感染HCoV-229E,在33°C下孵育2小时,洗涤后,在含有指定浓度DMSO或4E2RCat的培养基中孵育至多48小时。使用针对HCoV-229E S蛋白的单克隆抗体5-11H.6,以及辣根过氧化物酶标记的二抗,通过免疫过氧化物酶法定量病毒感染滴度(细胞内和细胞外),并用二氨基联苯胺和过氧化氢进行显色。采用 Karber 法计算滴度 [1] - 病毒 S 蛋白免疫荧光:将感染细胞固定,并用小鼠 IgG1 单克隆抗体 5-11H.6 染色,随后用 AlexaFluor-488 标记的抗小鼠山羊抗体(绿色)染色,最后用 DAPI(蓝色)染色细胞核。测定 S 蛋白阳性细胞的百分比 [1] - 脊髓灰质炎病毒感染:将 HeLa 细胞(6 孔板,每孔 3×10^5 个细胞)用 2 PFU/细胞的 1 型脊髓灰质炎病毒 Mahoney 株在室温下感染 30 分钟,洗涤后,用载体(1% DMSO)或 50 μM 4E2RCat 在 37°C 下孵育 4 小时。最后 30 分钟加入 [35S]甲硫氨酸(150 μCi/ml)。细胞裂解后,采用SDS-PAGE和放射自显影法进行分析[1] |
| 毒性/毒理 (Toxicokinetics/TK) |
在 12.5 μM 4E2RCat 浓度下,L132 细胞在处理 72 小时内未观察到细胞死亡/凋亡的诱导,这通过 Annexin V/碘化丙啶染色和流式细胞术检测得出(以 DMSO 对照组为基准,凋亡细胞比例设定为 1,未见增加)[1]。
- 在显著降低宿主蛋白合成的浓度下(例如 50 μM),4E2RCat 对脊髓灰质炎病毒复制没有影响,表明其在细胞水平上没有诱导非特异性抗病毒状态[1]。 |
| 参考文献 | |
| 其他信息 |
4E2RCat 是一种有机分子实体。
冠状病毒(HCoV-229E)的复制需要亚基因组 mRNA 的帽依赖性翻译。4E-BP1(可螯合 eIF4E)的过表达会损害冠状病毒的复制,这表明 eIF4F 是一个脆弱点。4E2RCat 可阻断 eIF4E-eIF4G 的相互作用,从而抑制冠状病毒的复制[1] - 4E2RCat 在降低冠状病毒感染性病毒产量方面比 4E1RCat(另一种 eIF4E-eIF4G 相互作用抑制剂)更有效,尽管两者在 TR-FRET 检测中的 IC50 值相似。在该系统中,4E2RCat 的效力似乎也高于 eIF4A 抑制剂 hippuristanol 和 silvestrol[1] - 计算机模拟预测,4E2RCat 可与 eIF4E 的 eIF4G/4E-BP 结合区域中的五个热点中的四个结合。将该化合物延伸至第五个未被占据的热点区域(图 3 中的品红色区域)可以提高亲和力 [1] - 该化合物是从分子库筛选中心网络对 217,341 个化合物进行超高通量筛选中发现的 [1] |
| 分子式 |
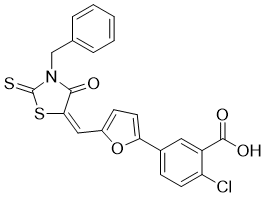
C22H14CLNO4S2
|
|---|---|
| 分子量 |
455.93
|
| 精确质量 |
455.005
|
| CAS号 |
432499-63-3
|
| PubChem CID |
2287238
|
| 外观&性状 |
Light yellow to yellow solid powder
|
| 密度 |
1.6±0.1 g/cm3
|
| 沸点 |
659.0±65.0 °C at 760 mmHg
|
| 闪点 |
352.3±34.3 °C
|
| 蒸汽压 |
0.0±2.1 mmHg at 25°C
|
| 折射率 |
1.760
|
| LogP |
5.09
|
| tPSA |
128
|
| 氢键供体(HBD)数目 |
1
|
| 氢键受体(HBA)数目 |
6
|
| 可旋转键数目(RBC) |
5
|
| 重原子数目 |
30
|
| 分子复杂度/Complexity |
724
|
| 定义原子立体中心数目 |
0
|
| SMILES |
C1=CC=C(C=C1)CN2C(=O)/C(=C\C3=CC=C(O3)C4=CC(=C(C=C4)Cl)C(=O)O)/SC2=S
|
| InChi Key |
WOBPZFKXPCYOLU-YBFXNURJSA-N
|
| InChi Code |
InChI=1S/C22H14ClNO4S2/c23-17-8-6-14(10-16(17)21(26)27)18-9-7-15(28-18)11-19-20(25)24(22(29)30-19)12-13-4-2-1-3-5-13/h1-11H,12H2,(H,26,27)/b19-11+
|
| 化学名 |
5-[5-[(E)-(3-Benzyl-4-oxo-2-sulfanylidene-1,3-thiazolidin-5-ylidene)methyl]furan-2-yl]-2-chlorobenzoic acid
|
| 别名 |
4E2R Cat 4E2RCat4E2R-Cat
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO : ~23.33 mg/mL (~51.17 mM)
|
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: 2.33 mg/mL (5.11 mM) in 10% DMSO + 40% PEG300 +5% Tween-80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 悬浮液;超声助溶。
例如,若需制备1 mL的工作液,可将100 μL 23.3 mg/mL澄清的DMSO储备液加入到400 μL PEG300中,混匀;再向上述溶液中加入50 μL Tween-80 +,混匀;然后加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.1933 mL | 10.9666 mL | 21.9332 mL | |
| 5 mM | 0.4387 mL | 2.1933 mL | 4.3866 mL | |
| 10 mM | 0.2193 mL | 1.0967 mL | 2.1933 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
|
|
|
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1


































 463611831
463611831
