| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 1mg |
|
||
| 5mg |
|
||
| 10mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| Other Sizes |
|

描述: 5-BDBD 是一种新型强效的 P2X4 受体拮抗剂。它能阻断中国仓鼠卵巢细胞中 P2X4 介导的电流,IC50 值为 0.50 μM。
| 靶点 |
P2X4 Receptor
P2X4 receptor (P2X4R) [1] P2X4 receptor (P2X4R) [2] |
|---|---|
| 体外研究 (In Vitro) |
5-BDBD 的 IC50 为 0.75 μM,它以浓度依赖的方式抑制表达 rP2X4R 的 HEK293 细胞中 10 μM ATP 产生的电流 [1]。5-BDBD 使 EC50 从 4.7 μM 增加到 15.9 μM,从而使 ATP 浓度-反应曲线右移 [1]。P2X1R、P2X2aR、P2X2bR、P2X3R、P2X4R 和 P2X7R 均可使用 5-BDBD 进行区分 [1]。P2X 受体 (P2XR) 是一类 ATP 门控离子通道,表达于多种可兴奋和非可兴奋细胞中。尽管过去几十年对这些受体的结构和功能的研究取得了巨大进展,但目前仍然缺乏针对 P2XR 亚型(尤其是 P2X4R)的特异性强效拮抗剂。本研究详细探讨了P2X4R拮抗剂5-(3-溴苯基)-1,3-二氢-2H-苯并呋喃并[3,2-e]-1,4-二氮杂卓-2-酮(5-BDBD)对大鼠P2X4R介导的ATP诱导电流的影响,并比较了其对其他大鼠P2XR的特异性。研究发现,5-BDBD是一种强效的P2X4R拮抗剂,在ATP刺激前2分钟及刺激期间应用时,其IC50值为0.75 μM。此外,在10 μM浓度下,5-BDBD不影响ATP诱导的P2X2aR、P2X2bR和P2X7R电流幅度或受体脱敏模式。然而,浓度为 10 μM 的 5-BDBD 可分别抑制 P2X1R 和 P2X3R 门控电流 13% 和 35%,而浓度为 0.75 μM 的 5-BDBD 则无此作用。此外,研究人员在对大鼠海马切片进行的长时程增强 (LTP) 实验中研究了 5-BDBD 的作用,发现该拮抗剂可部分降低 LTP,而这种反应被认为部分由内源性 P2X4R 介导。这些结果表明,5-BDBD 可用于研究中枢神经系统中 P2X4R 的内源性效应,并且当 P2X4R 和其他 P2XR 在同一组织中共表达时,该拮抗剂可以区分它们[1]。
在 HEK293 细胞中表达的大鼠 P2X4R (rP2X4R) 上,5-BDBD 以浓度依赖性方式抑制 ATP (10 µM) 诱导的电流,当预先施加 2 分钟并与 ATP 一起施加时,IC50 为 0.75 ± 0.27 µM。用 10 µM 5-BDBD 洗脱 5 分钟后,电流几乎完全恢复 [1]。 预先施加 1 µM 5-BDBD 2 分钟后,rP2X4R 的 ATP 浓度-反应曲线右移,ATP 的 EC50 值从 4.7 ± 1.8 µM 增加到 15.9 ± 3.9 µM,但最大反应没有显著改变(对照组:2.5 ± 0.6 nA,5-BDBD 处理组:2.0 ± 0.6 nA)[1]。 在 10 µM 浓度下,5-BDBD 不影响表达大鼠 P2X2aR、P2X2bR 或 1 mM ATP 诱导的 HEK293 细胞中电流的峰值幅度或脱敏模式(P2X2a/bR 为 10 µM,P2X7R 为 1 mM)。 P2X7R。在相同浓度下,该化合物对P2X3R门控电流产生适度但显著的抑制作用(抑制率为35.9 ± 10.1%),对P2X1R门控电流产生较小但不显著的抑制作用(抑制率为12.7 ± 5.6%),而相同条件下对P2X4R的抑制率为83.2 ± 3.1% [1]。 蛋白质印迹分析证实了大鼠海马突触和突触外区域存在P2X4R的表达,但未检测到P2X1R或P2X3R的表达[1]。 在慢性偏头痛小鼠模型中,重复注射硝酸甘油(NTG)(10 mg/kg,腹腔注射,每隔一天一次,持续9天)导致三叉神经尾核(TNC)中P2X4R蛋白表达随时间增加,最早在第3天即可检测到,并在第9天检测到P2X4R mRNA表达增加。 [2]. 对这些小鼠的三叉神经脊束核(TNC)进行双重免疫荧光标记显示,几乎所有P2X4R阳性细胞均与Iba1(一种小胶质细胞标记物)共标记,表明小胶质细胞表达P2X4R[2]. 在反复注射硝酸甘油(NTG)的小鼠的TNC中,5-BDBD治疗(28 mg/kg,腹腔注射,每次NTG注射前)显著降低了NTG诱导的c-Fos阳性细胞的增加(从84.9 ± 11.8个细胞/切片降至34.1 ± 8.7个细胞/切片),并减弱了NTG诱导的降钙素基因相关肽(CGRP)免疫反应性纤维光密度的降低[2]. |
| 体内研究 (In Vivo) |
使用 5-BDBD(28 mg/kg;腹腔注射;每日一次,连续 9 天)可以完全模拟反复注射硝酸甘油 (NTG) 诱发的基础痛觉过敏 [2]。研究人员使用了一种反复间歇性注射硝酸甘油 (NTG) 的动物模型,该模型与慢性偏头痛 (CM) 非常相似。他们使用 von Frey 纤维丝试验评估了 NTG 诱发的基础和急性机械性痛觉过敏。然后,研究人员检测了三叉神经尾核 (TNC) 中的 Iba1 免疫反应性 (Iba1-IR) 和 P2X4R 表达。为了解小胶质细胞和 P2X4R 对 CM 中枢敏化的影响,研究人员检测了小胶质细胞活化抑制剂米诺环素和 P2X4R 拮抗剂 5-BDBD 是否会改变 NTG 诱发的机械性痛觉过敏。此外,研究人员还评估了5-BDBD对三叉神经核(TNC)内c-Fos和降钙素基因相关肽(CGRP)表达的影响。结果:慢性间歇性给予硝酸甘油(NTG)导致急性及慢性基础机械性痛觉过敏,并伴有小胶质细胞活化和P2X4受体表达上调。米诺环素显著降低了基础痛觉过敏,但并未改变NTG诱导的急性痛觉过敏。米诺环素还降低了小胶质细胞活化。5-BDBD完全阻断了NTG诱导的基础及急性痛觉过敏。这种作用与显著抑制NTG诱导的TNC内c-Fos蛋白和CGRP释放的增加有关。结论:我们的结果表明,抑制小胶质细胞活化可能有助于预防偏头痛慢性化。此外,研究人员推测 P2X4R 可能与 TNC 中的小胶质细胞-神经元信号有关,这有助于 CM 的中枢敏化 [2]。
在大鼠海马切片中,用 10 µM 5-BDBD 预孵育 60 分钟可显著降低长期增强 (LTP) 的晚期,这反映在 LTP 诱导后 40-60 分钟测量的场兴奋性突触后电位 (fEPSP) 斜率值的显著降低。相反,在相同条件下,非特异性拮抗剂PPADS (3 µM) 和P2X3R拮抗剂AF-353 (100 nM) 对LTP没有显著影响[1]。 在慢性偏头痛小鼠模型中(重复注射NTG,10 mg/kg,腹腔注射,每隔一天一次,持续9天),预先注射5-BDBD(28 mg/kg,腹腔注射,在每次NTG/生理盐水注射前立即注射)可完全阻断慢性基础机械性痛觉过敏和急性NTG诱导的机械性痛觉过敏(通过von Frey纤维测试测量)[2]。 在同一小鼠模型中,预先注射5-BDBD(28 mg/kg,腹腔注射,在每次NTG注射前注射)可显著抑制NTG诱导的三叉神经尾核(TNC)中c-Fos蛋白表达和CGRP释放的增加[2]。 |
| 细胞实验 |
受体转染和电流测量。[1]
实验采用大鼠P2X受体。HEK293细胞常规培养于含10% (v/v)胎牛血清和100 µg/ml庆大霉素的DMEM培养基中。细胞以每35 mm培养皿50万个细胞的密度接种。接种24小时后进行瞬时转染,使用2 µg DNA和5 µl LipofectAMINE 2000试剂,溶于2 ml无血清Opti-MEM培养基中。孵育4.5小时后,将转染混合物更换为正常培养基。转染后24-48小时进行实验。记录前,将转染细胞机械分散,并在35 mm培养皿中重新培养2-10小时。在室温下,使用全细胞膜片钳记录技术对细胞进行电生理实验。使用 Axopatch 200B 膜片钳放大器记录电流,并使用 2 kHz 低通贝塞尔滤波器进行滤波。膜片钳电极由硼硅酸盐玻璃制成,采用 Flaming Brown 水平拉丝机拉制,并经热抛光处理,最终尖端电阻为 2–4 MΩ。所有电流记录均使用 pClamp 9 软件和 Digidata 1322A 模数转换器进行采集和存储。膜片钳电极内填充溶液成分如下(单位:mM):142 NaCl、1 MgCl₂、10 EGTA 和 10 HEPES,用 10 M NaOH 调节 pH 至 7.35。内液渗透压为 306 mOsm。浴液成分如下(单位:mM):142 NaCl、3 KCl、1 MgCl₂、2 CaCl₂、10 葡萄糖和 10 HEPES,用 10 M NaOH 调节 pH 至 7.35。该溶液的渗透压为 295–305 mOsm。ATP 每日用浴液缓冲液配制,并使用快速换液系统进行施加。5-BDBD 储备液用二甲基亚砜配制,并分装后储存于 –20 °C。电流反应记录于钳制在 –60 mV 的单细胞。浓度-反应数据通过记录施加于单细胞的不同 ATP 浓度下的电流获得,每次施加之间间隔 1–5 分钟的洗脱期,并将结果归一化至最高电流幅度。电生理记录使用表达大鼠 P2X 受体的 HEK293 细胞。将细胞以每35 mm培养皿50万个的密度接种,24小时后,使用2 µg DNA和5 µl脂质体转染试剂在无血清培养基中进行瞬时转染。4.5小时后,将转染混合物更换为正常培养基。转染后24-48小时进行实验。记录前,将转染细胞机械分散并重新培养2-10小时。在室温下进行全细胞膜片钳记录。细胞钳制在-60 mV。使用快速换液系统施加ATP。5-BDBD的储备液用DMSO配制[1]。 P2X4R表达分析使用HEK293细胞(转染和未转染)和大鼠脑组织。在含有蛋白酶和磷酸酶抑制剂混合物的缓冲液中获得蛋白质提取物。将蛋白质进行SDS-PAGE电泳,然后转移至硝酸纤维素膜。膜用5%脱脂奶粉封闭,然后在4℃下与P2X4R一抗孵育过夜,随后与二抗孵育。使用化学发光底物检测免疫反应[1]。 |
| 动物实验 |
动物/疾病模型:雄性C57BL/6小鼠[2]
剂量:28 mg/kg 给药途径:腹腔注射;每日一次,连续9天 实验结果:预防硝酸甘油诱导的机械性过敏。 海马切片制备。[1] 断头后,将脑组织迅速浸入冰冷的解剖缓冲液中,该缓冲液含有(单位:mM):212.7蔗糖;5 KCl;1.25 NaH2PO4;3 MgSO4;1 CaCl2;26 NaHCO3;以及10葡萄糖;pH 7.4。解剖海马,并使用振动切片机从中间三分之一处获得横切片(厚度为400 µm)。将脑片转移至含有饱和95% O₂/5% CO₂的人工脑脊液(aCSF)的界面储存室,并在37℃下至少放置1小时。之后,将脑片置于aCSF中,或在记录前分别用5-BDBD、PPADS或AF-353孵育1小时。aCSF的成分(单位:mM)为:124 NaCl;5 K₁;1.25 NaH₂PO₄;1.0 MgCl₂;2.0 CaCl₂;26 NaHCO₃;10葡萄糖;pH 7.4。随后将单个脑片转移至记录室,使其完全浸没在aCSF中,并持续灌注(2 ml/min)。 长时程增强(LTP)实验。 [1] 使用连接至刺激隔离器的双极电极,在Schaffer侧支通路处,每隔15秒对对照组和5-BDBD孵育的海马进行电刺激(双相恒流刺激,200 μs),并在放射层记录电刺激信号,以观察CA1区场突触后电位(fEPSP)。记录电极为充满人工脑脊液(ACSF)的玻璃微电极(1–2 MΩ)。在每次实验开始时,通过逐渐增加刺激强度,直至达到最大反应的50%,绘制刺激-反应曲线。在20分钟的稳定基线期后,通过θ节律刺激(TBS)诱发长时程增强(LTP)。TBS包含5组刺激,组间间隔为10秒。每个刺激序列包含10个5 Hz的脉冲串,每个脉冲串包含4个100 Hz的脉冲。TBS刺激后,数据采集持续1小时。数据采集使用细胞外放大器和数据采集板(美国国家仪器公司,德克萨斯州奥斯汀),并通过Igor Pro软件进行控制。LTP值通过将fEPSP归一化,并将基线值设为100%来构建。LTP的幅度在TBS刺激后40至60分钟之间观察到。为了解小胶质细胞和P2X4R对NTG诱导的痛觉过敏的影响,我们用小胶质细胞抑制剂米诺环素或P2X4R选择性拮抗剂5-BDBD处理动物。米诺环素和5-BDBD均溶解于DMSO溶液中,DMSO溶液用作溶剂对照。在注射硝酸甘油/生理盐水之前,立即给予米诺环素/载体或5-BDBD/载体[14, 15]。测试日小鼠的处理方法如下:小鼠适应测试室后,15-20分钟后测量机械阈值基线值,然后腹腔注射米诺环素(30 mg/kg)、5-BDBD(28 mg/kg)或载体;之后注射硝酸甘油/生理盐水,2小时后测试急性机械反应。此过程每隔一天重复一次,持续9天。[2] 在海马长时程增强(LTP)实验中,用氟烷麻醉一个月大的雄性Sprague-Dawley大鼠并断头处死。迅速取出脑组织并浸入冰冷的解剖缓冲液中。解剖海马体,并使用振动切片机获得400 µm厚的横切片。将切片转移至含有饱和95% O₂/5% CO₂的人工脑脊液(aCSF)的界面储存室中,并在37°C下至少放置1小时。之后,将切片继续置于aCSF中,或在记录前分别用10 µM 5-BDBD、3 µM PPADS或100 nM AF-353孵育1小时。为了诱导长时程增强(LTP),每隔15秒对Schaffer侧支通路进行电刺激以诱发场电位。在20分钟的稳定基线期后,通过θ节律刺激(TBS)诱发LTP,该刺激由5组间隔10秒的刺激组成。每个刺激序列包含10个5 Hz的脉冲串,每个脉冲串包含4个100 Hz的脉冲。数据采集在TBS刺激后持续1小时[1]。 慢性偏头痛模型采用雄性C57BL/6小鼠(8-10周龄,18-20 g)。小鼠每隔一天腹腔注射10 mg/kg硝酸甘油(NTG)或溶剂,持续9天(第1、3、5、7、9天)。为了研究P2X4R阻断的作用,在注射NTG/生理盐水前,立即用P2X4R拮抗剂5-BDBD或其溶剂(DMSO溶液)处理小鼠。 5-BDBD 的给药方案为 28 mg/kg,腹腔注射。测试当日的流程如下:首先测量机械阈值基线值,然后给予 5-BDBD/载体,接着注射 NTG/生理盐水,2 小时后测试急性机械反应。对于慢性基础痛觉过敏,每次注射 NTG 前均测试机械阈值。采用 von Frey 纤维丝,根据上下法 [2] 测试后爪足底的机械敏感性。 |
| 参考文献 | |
| 其他信息 |
总之,我们已鉴定出5-BDBD是一种特异性的P2X4R拮抗剂。此类分子的开发将有助于阐明P2XR在生理过程中的作用,并有助于设计针对涉及这些受体的疾病的特异性药物。[1] 在我们的研究中,CGRP免疫反应性纤维主要分布于三叉神经核(TNC)浅层,该区域主要与伤害性信息的处理相关。5-BDBD预处理可减轻NTG诱导的TNC中CGRP免疫反应性的变化,从而调节三叉神经系统的激活。P2X4R诱导CGRP释放的细胞机制可能与脑源性神经营养因子(BDNF)有关。既往研究表明,ATP刺激的小胶质细胞可通过P2X4R信号通路在体外和体内释放BDNF。此外,BDNF可通过TrkB受体调节感觉神经元中CGRP的表达和释放。慢性偏头痛的发生与三叉神经感觉通路的中枢敏化有关,这一过程涉及小胶质细胞活化和P2X4R表达增加。抑制小胶质细胞可能有效预防偏头痛的慢性化,但对终止急性偏头痛发作无效。此外,我们假设P2X4R可能参与三叉神经核(TNC)中的小胶质细胞-神经元信号传导。抑制P2X4R功能可能是治疗慢性偏头痛的一种潜在方法。 [2] 5-BDBD(5-(3-溴苯基)-1,3-二氢-2H-苯并呋喃并[3,2-e]-1,4-二氮杂卓-2-酮)是一种苯二氮卓类衍生物,最初被描述为竞争性正位拮抗剂,但最近的放射性配体研究表明其作用机制为变构作用。它是一种强效的P2X4R拮抗剂,可用于研究中枢神经系统中内源性P2X4R介导的功能,因为它能够区分在同一组织中共表达的P2X4R和其他P2XR亚型(P2X1、P2X2a、P2X2b、P2X3、P2X7)。 P2X4R 的优势之一是它对苏拉明和 PPADS 等广谱嘌呤能拮抗剂具有抵抗性 [1]。
在慢性偏头痛小鼠模型中,研究表明 P2X4R 拮抗剂 5-BDBD 可能参与三叉神经尾核 (TNC) 的小胶质细胞-神经元信号传导,抑制 P2X4R 功能可能是治疗慢性偏头痛的潜在方法 [2]。 |
| 分子式 |
C17H11BRN2O2
|
|---|---|
| 分子量 |
355.1854
|
| 精确质量 |
354
|
| 元素分析 |
C, 57.49; H, 3.12; Br, 22.50; N, 7.89; O, 9.01
|
| CAS号 |
768404-03-1
|
| PubChem CID |
9841560
|
| 外观&性状 |
White to light yellow solid powder
|
| LogP |
3.505
|
| tPSA |
58.09
|
| 氢键供体(HBD)数目 |
1
|
| 氢键受体(HBA)数目 |
3
|
| 可旋转键数目(RBC) |
1
|
| 重原子数目 |
22
|
| 分子复杂度/Complexity |
481
|
| 定义原子立体中心数目 |
0
|
| SMILES |
BrC1=C([H])C([H])=C([H])C(=C1[H])C1C2=C(C3=C([H])C([H])=C([H])C([H])=C3O2)N([H])C(C([H])([H])N=1)=O
|
| InChi Key |
NKYMVQPXXTZHSF-UHFFFAOYSA-N
|
| InChi Code |
InChI=1S/C17H11BrN2O2/c18-11-5-3-4-10(8-11)15-17-16(20-14(21)9-19-15)12-6-1-2-7-13(12)22-17/h1-8H,9H2,(H,20,21)
|
| 化学名 |
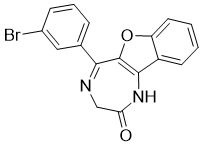
5-(3-bromophenyl)-1,3-dihydro-[1]benzofuro[3,2-e][1,4]diazepin-2-one
|
| 别名 |
5-BDBD; 768404-03-1; 5-(3-Bromophenyl)-1,3-dihydro-2H-benzofuro[3,2-e]-1,4-diazepin-2-one; 5 BDBD;5BDBD; 5-(3-bromophenyl)-1,3-dihydro-[1]benzofuro[3,2-e][1,4]diazepin-2-one; CHEMBL2180179; DTXSID30431707; 5-(3-bromophenyl)-1,3-dihydro-(1)benzofuro(3,2-e)(1,4)diazepin-2-one;
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO : ~62.5 mg/mL (~175.96 mM)
|
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.08 mg/mL (5.86 mM) (饱和度未知) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 20.8 mg/mL澄清DMSO储备液加入400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: ≥ 2.08 mg/mL (5.86 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 例如,若需制备1 mL的工作液,可将 100 μL 20.8 mg/mL 澄清 DMSO 储备液加入到 900 μL 玉米油中并混合均匀。 View More
配方 3 中的溶解度: 9.8 mg/mL (27.59 mM) in 15% Solutol HS 15 10% Cremophor EL 35% PEG 400 40% water (这些助溶剂从左到右依次添加,逐一添加), 悬浊液; 超声助溶。 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.8154 mL | 14.0770 mL | 28.1539 mL | |
| 5 mM | 0.5631 mL | 2.8154 mL | 5.6308 mL | |
| 10 mM | 0.2815 mL | 1.4077 mL | 2.8154 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
|
|
|
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1


































 463611831
463611831
