| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| Other Sizes |
|
| 靶点 |
mGluR2 ( EC50 = 285 nM )
AZD-8529 mesylate increases the effects of glutamate at mGluR2 with an EC50 of 195 nM [1]. AZD-8529 mesylate does not cause antagonist reactions on mGluRs at 25 μM[1]. AZD8529 potentiation of mGluR2 receptor function[1] We assessed the effect of AZD8529 at the human mGluR2 receptor by measuring the potentiation of [35S]GTPγS binding in the presence of increasing concentrations of exogenously applied agonist (L-glutamate). AZD8529 potentiated the effects of glutamate at mGluR2 with an EC50 of 195±62 nM and an Emax of 110%±11% (Figure 1A). In order to assess the selectivity of AZD8529 against the family of mGluRs, we used fluorescence-based assays. AZD8529 potentiated mGluR2 activity with an EC50 of 285±20 nM and did not produce any positive allosteric modulator responses at 20-25 M on the mGluR1, 3, 4, 5, 6, 7, and 8 subtypes (Table 1). In addition, at 25 μM AZD8529 did not elicit antagonist responses on mGluRs. When AZD8529 (10 μM) was studied in a broad receptor screen (Table 2), we observed >50% inhibition of ligand biding at adenosine A3 receptors (51% inhibition) and the norepinephrine transporter (NET, IC50=4.73 μM). In vitro electrophysiology tests [2] The mGlu2/3 receptor agonist DCG-IV induced inhibitory effects on fEPSPs in the pyramidal cells in the CA1 region of the hippocampus. The inhibitory effects of DCG-IV on synaptic transmission were potentiated by AZD8418 and AZD8529, with EC50 values of 0.86 and 1.4 μM, respectively (Fig. 2a). AZD8418 produced a greater maximal potentiation (79.9 ± 4.4) than AZD8529 (61.4 ± 4.3, p < 0.05). The mGlu2/3 receptor antagonist LY341495 (1 μM) effectively blocked the inhibition of fEPSPs that was induced by DCG-IV alone (10 nM) or the combination of DCG-IV with either AZD8529 (10 μM) or AZD8418 (10 μM; Fig. 2b). |
|---|---|
| 体外研究 (In Vitro) |
AZD-8529 mesylate 增强谷氨酸对 mGluR2 的作用,EC50 为 195 nM [1]。 AZD-8529 甲磺酸盐在 25 μM 时不会引起 mGluR 拮抗剂反应[1]。
AZD8529增强mGluR2受体功能[1] 我们通过测量在外源性激动剂(L-谷氨酸)浓度增加的情况下[35S]GTPγS结合的增强,评估了AZD8529对人类mGluR2受体的影响。AZD8529增强了谷氨酸对mGluR2的影响,EC50为195±62 nM,Emax为110%±11%(图1A)。为了评估AZD8529对mGluRs家族的选择性,我们使用了基于荧光的检测方法。AZD8529增强mGluR2活性,EC50为285±20 nM,在20-25 M下对mGluR1、3、4、5、6、7和8亚型没有产生任何正变构调节剂反应(表1)。此外,在25μM时,AZD8529对mGluRs没有引起拮抗剂反应。当在广泛的受体筛选中研究AZD8529(10μM)时(表2),我们观察到配体结合腺苷A3受体(51%抑制)和去甲肾上腺素转运蛋白(NET,IC50=4.73μM)的抑制率>50%。 体外电生理试验[2] mGlu2/3受体激动剂DCG-IV对海马CA1区锥体细胞中的fEPSPs具有抑制作用。AZD8418和AZD8529增强了DCG-IV对突触传递的抑制作用,EC50值分别为0.86和1.4μM(图2a)。AZD8418产生的最大增强作用(79.9±4.4)大于AZD8529(61.4±4.3,mGlu2/3受体拮抗剂LY341495(1μM)有效阻断了DCG-IV单独(10 nM)或DCG-IV与AZD8529(10μM)或AZD8418(10μM)联合诱导的fEPSPs的抑制作用;图2b)。 在表达人mGluR2的CHO细胞膜进行的[35S]GTPγS结合实验中,AZD8529 增强了激动剂(L-谷氨酸)诱导的受体活化(EC50 = 195 ± 62 nM, Emax = 110% ± 11%)。[1] 在使用表达嵌合人mGluR受体的HEK-293细胞进行的荧光实验中,AZD8529 是mGluR2的正向变构调节剂(EC50 = 285 ± 20 nM, Emax = 59.9% ± 14%),但对mGluR1、3、4、5、6、7或8无显著影响。[1] 在食蟹猴脑切片上使用[35S]GTPγS放射自显影技术,AZD8529 (3 µM) 显著增强了亚最佳浓度的mGluR2/3激动剂LY379268 (1 µM) 在皮层、海马和纹状体诱导的信号。这种增强可被mGluR2/3拮抗剂LY341495 (1 µM) 逆转。[1] |
| 体内研究 (In Vivo) |
AZD8529(0.3 mg/kg,肌肉注射)可减少松鼠猴中尼古丁启动诱导和提示诱导的恢复[1]。 AZD-8529(30 mg/kg;腹腔注射)可降低自由活动大鼠伏隔核中尼古丁诱导的细胞外多巴胺增加[1]。动物模型:Sprague-Dawley大鼠[1] 剂量:10 mg/kg、30 mg/kg 给药方式:腹腔注射;尼古丁注射前 2 小时 结果:减少了自由活动大鼠伏隔核中由尼古丁(0.4 mg/kg,皮下)引起的细胞外多巴胺增加 [1]。
在猴子中,AZD8529在不影响食物自我给药的剂量(.3-3mg/kg)下减少了尼古丁的自我给药。AZD8529还减少了尼古丁引发和线索诱导的尼古丁恢复,在药物消失后寻求增强反应。在大鼠中,AZD8529减少了尼古丁诱导的伏隔核多巴胺释放。 结论:这些结果为mGluR2的正变构调节剂在尼古丁强化和复发的非人灵长类模型中的有效性提供了证据。这类药物应考虑用于尼古丁成瘾治疗。 AZD8418(0.37、1.12、3.73、7.46和14.92 mg/kg)和AZD8529(1.75、5.83、17.5和58.3 mg/kg)的急性治疗抑制了尼古丁的自我给药,对食物维持反应没有影响。AZD8418的慢性治疗减弱了尼古丁的自我给药,但对这种作用的耐受性迅速发展。慢性AZD8529给药对尼古丁自我给药的抑制作用在整个14天的治疗过程中持续存在。任何一种PAM的慢性治疗都会抑制食物的自我给药。AZD8418(急性)和AZD8529(急性和亚慢性)阻断了线索诱导的尼古丁和食物寻求行为的恢复。结论:这些发现表明,mGlu2受体在自我给药尼古丁的增强特性以及之前与尼古丁给药相关的线索的动机影响(即线索诱导的尼古丁寻求行为的恢复)中起着重要作用。因此,mGlu2-PAM可能是帮助人们戒烟和防止复发的有用药物[2]。 在训练为静脉自我给予尼古丁(30 µg/kg/次)的松鼠猴中,预处理 AZD8529 (0.3、1、3 和 10 mg/kg, 肌注, session前3小时) 在连续三个测试session中显著降低了尼古丁自我给药,且未观察到耐受性。0.3-3 mg/kg的剂量不影响食物自我给药,而更高剂量(10 和 30 mg/kg)则产生影响。[1] AZD8529 (0.3、1 和 3 mg/kg, 肌注) 剂量依赖性地降低了松鼠猴中尼古丁引燃诱导(0.1 mg/kg, 静脉)和线索诱导的熄灭后尼古丁寻求行为的复吸。[1] 在大鼠中,AZD8529 (30 mg/kg, 腹腔注射,但10 mg/kg无效) 减弱了尼古丁(0.4 mg/kg, 皮下注射)诱导的伏隔核壳区细胞外多巴胺水平的升高。AZD8529 单独给药对多巴胺水平无影响。[1] |
| 酶活实验 |
功能性mGluR2测定[1]
受体选择性测定[1] 为了确定mGluR家族中AZD8529的选择性,我们使用了基于荧光的检测和表达人mGluR构建体的HEK 293细胞系。所述细胞系表达嵌合融合构建体hmGluR2/hCaR*、hmGluR1/hCaR+、hmGluR3/hCaR+hmGluR4/hCaR*hmGluR5/hCaR>hmGluR6/hCaR=hmGluR7/hCaR*hmGluR8/hCaR*,各自包含人mGluR的细胞外结构域和跨膜结构域,以及如前所述与混杂嵌合蛋白Gqi5融合的人钙受体的细胞内结构域。 受体筛查[1] 我们根据已发表的方法,使用放射性配体结合分析(MDS Pharma)评估了10μM下AZD8529的脱靶效应。我们为每种测定都运行了参考标准。我们使用数据分析工具箱(MDL信息系统)的非线性最小二乘回归分析来确定IC50值。 [35S]GTPγS结合人mGlu2-CHO膜[1] 我们使用由表达人mGluR2的CHO细胞系制备的膜,并以闪烁邻近试验(SPA)形式进行了试验。我们将表达人mGluR2的中国仓鼠卵巢(CHO)细胞培养至约80%融合,在冰冷的磷酸盐缓冲盐水中洗涤细胞,并冷冻保存直至膜制备。测定缓冲液含有0.05 M HEPES、0.10 M NaCl、0.01 M MgCl2,pH 7.4,加上100 M二硫苏糖醇和3 M鸟苷二磷酸。我们通过在含有AZD8529或载体的测定缓冲液中加入小麦胚芽凝集素SPA珠(0.75mg/ml)和膜(6g/ml)的混合物来开始测定。孵育15分钟后,我们加入了一种含有[35S]GTPγS和L-谷氨酸盐的溶液(最终浓度为100 pM[35S]GTPγS和0-100 M谷氨酸盐)。在室温下孵育(60分钟)后,我们对测定板进行离心,并在TopCount™闪烁计数器上读取。我们分别测定了在没有谷氨酸和存在100-M谷氨酸的情况下[35S]GTPγS结合的0%和100%水平。我们根据AZD8529的浓度反应曲线估计了AZD8529s对mGluR2激活的调节活性,并用4参数逻辑斯谛方程进行了拟合,以计算表观效力(EC50)和最大效力(Emax)。 使用从表达人mGluR2的CHO细胞系制备的膜,以闪烁亲近测定法(SPA)形式评估 AZD8529 对mGluR2功能的影响。测定缓冲液含有HEPES、NaCl、MgCl2、二硫苏糖醇和鸟苷二磷酸。将含有 AZD8529 或溶媒的测定缓冲液中的小麦胚芽凝集素SPA珠和膜混合物孵育15分钟。然后加入含有[35S]GTPγS和不同浓度L-谷氨酸的溶液。在室温下孵育60分钟后,离心平板并在闪烁计数仪上读数。将不含谷氨酸和含最大浓度谷氨酸(100 µM)时的[35S]GTPγS结合分别定义为0%和100%活性。拟合 AZD8529 的浓度-反应曲线以计算EC50和Emax。[1] 使用标准放射性配体结合实验,在单一浓度(10 µM)的 AZD8529 下,针对一组受体、离子通道和转运体进行了脱靶效应筛选。每个实验都运行了参考标准品。[1] |
| 细胞实验 |
[35S]食蟹猴脑片GTPγS放射自显影术[1]
我们用戊巴比妥钠(100mg/kg)麻醉猴子,用生理盐水灌注,然后取出大脑并将其冷冻在冷却的异戊烷中。我们在低温恒温器上切下20-μm的纹状体和海马切片,将切片安装在载玻片上,并在80°C下储存直至使用。在实验当天,我们在真空室中将切片加热到室温超过3小时。我们在50 mM Tris-HCl、3 mM MgCl2、0.2 mM EGTA、100 mM NaCl和0.2 mM DTT(Tris测定缓冲液,TAB)中孵育切片;pH 7.4,在25°C下孵育10分钟。然后,我们将载玻片在含有2 mM二磷酸鸟苷(GDP)的TAB中,在25℃下孵育15分钟。我们将载玻片置于以下四种条件之一中,在25°C下放置2小时:基础:TAB+2 mM GDP+0.04 nM[35S]GTPγS;单独使用激动剂:TAB+2 mM GDP+0.04 nM[35S]GTPγS+1μM LY379268;单独使用调节剂:TAB+2 mM GDP+0.04 nM[35S]GTPγS+3μMAZD8529;调节剂+激动剂:TAB+2 mM GDP+0.04 nM[35S]GTPγS+1μM LY379268+3μM AZD8529;调节剂+激动剂+拮抗剂:TAB+2 mM GDP+0.04 nM[35S]GTPγS+1μM LY379268+3μM AZD8529+1μM LY341495。然后,我们在4°C 50 mM Tris-HCl(pH 7.4)中洗涤切片2次,每次5分钟,在冰冷的H2O中漂洗,并风干载玻片。然后,我们将载玻片暴露在Biomax MR胶片上2天,并使用标准技术进行显影、数字化和分析。 切片记录[2] 将切片浸入切片室中,以1-2ml/min的流速浸泡在32°C aCSF中。用铂丝制成的砝码将切片固定到位。用连接到隔离脉冲刺激器(型号2100)的单极钨电极(型号575300,0.5-1mΩ)刺激辐射层中的Schaeffer侧支纤维。用从硼硅酸盐玻璃(型号TW150-4)中拔出并填充2mM NaCl的电极记录CA1层锥体细胞的细胞外群体尖峰。用每30秒一次的10毫秒脉冲刺激切片。建立基线反应(最大值的50-70%),然后记录大约15分钟的控制期。在控制期后,将化合物浴敷40分钟或直至达到稳态响应。 |
| 动物实验 |
Sprague-Dawley 大鼠
10 mg/kg,30 mg/kg 腹腔注射;尼古丁注射前 2 小时 尼古丁自我给药[1] 此阶段持续 14 周,包括周一至周五的 1 小时实验。每次实验开始前,我们将猴子放在有机玻璃椅上,并用腰部锁将其固定在坐姿。我们首先训练猴子在固定比率强化程序(FR10,超时 60 秒)下按压杠杆,强化物为静脉注射尼古丁(30 μg/kg/次)。用 1 ml 生理盐水冲洗导管后,我们将其连接到电动注射器。每次实验开始时,白色照明灯关闭,绿色刺激灯打开;按压10次后,绿灯熄灭,琥珀灯亮起2秒,同时注射尼古丁(0.2毫升)。在60秒的暂停期内,实验箱处于黑暗状态,按压杠杆不会产生任何预设后果。当连续至少5次实验的反应变异性小于15%时,我们测试了AZD8529预处理(0.03、0.3、1、3和10毫克/千克,肌注,实验前3小时)对尼古丁自我给药的影响,共进行了3次实验;我们将这些数据与每次实验前连续3次注射赋形剂的预处理数据进行了比较。3小时的预处理时间是基于阿斯利康Tmax药代动力学研究(数据未显示)。 尼古丁渴求的恢复[1] 我们进行了为期9周的研究的这一阶段。我们首先测试了猴子在药物强化反应消退后,尼古丁启动诱导的复吸情况。然后,我们重新训练它们在5天内自我给药尼古丁,并在药物强化反应消退后测试了它们线索诱导的复吸情况。我们测试了3 mg/kg或更低剂量的AZD8529,因为3 mg/kg是能减少尼古丁自我给药但不减少食物自我给药的最高有效剂量。 尼古丁启动诱导的复吸[1] 在猴子进行每日消退训练后,我们测试了尼古丁启动诱导的复吸情况。在消退训练中,按压操纵杆会导致生理盐水输注以及之前与尼古丁输注配对的视觉线索,但不会出现尼古丁。在每次消退训练前,我们均给予非条件性生理盐水注射,作为尼古丁启动注射的载体对照。至少经过两次消退训练后,当反应达到较低且稳定的水平时,我们测定了预先注射AZD8529(0.3、1 或 3 mg/kg,肌注)或其载体对尼古丁(0.1 mg/kg,静脉注射)诱导的复吸的影响。我们在测试开始前立即给予尼古丁启动注射。测试期间,按压操作杆(FR10)仍然只产生生理盐水注射和离散提示。我们还测试了 3 mg/kg 的 AZD8529 对生理盐水启动的影响,以确定单独使用 AZD8529 是否会影响消退后的尼古丁渴求。 线索诱导的复吸 [1] 完成尼古丁启动测试后,我们对猴子进行了约 10 次的尼古丁自我给药训练。然后,我们进行了 3 次消退训练,期间按压杠杆没有强化后果(既没有尼古丁也没有线索);此外,在这些训练之前,我们没有给猴子注射生理盐水启动。消退训练结束后,我们确定了预先注射 AZD8529(0.3、1 或 3 mg/kg,肌注)或其溶剂对线索诱导的复吸的影响。在测试过程中,按压杠杆(FR10)会触发静脉输注生理盐水和呈现视觉提示。我们还测定了3 mg/kg AZD8529对无提示条件下消退反应的影响。每次提示诱导的恢复测试后,都会进行一到两次消退训练。 食物自我给药[1] 我们在另一组猴子中测定了AZD8529的作用。这些猴子在与尼古丁相同的强化程序条件下(FR 10,TO 60 s)自我给药190 mg的食物颗粒。我们限制了食物摄入量,以维持猴子的体重在有利于食物强化反应的水平(不超过自由摄食体重的10%)。本组每次实验中给予的强化物数量以及反应率与尼古丁组非常相似(图 2)。我们连续三次注射 AZD8529(3、10 和 30 mg/kg,肌注),每次注射前均进行三次载体注射。 AZD8529在松鼠猴体内的血浆浓度[1] 为了确定行为实验期间的血浆浓度是否达到人类可耐受的水平(根据阿斯利康公司提供的信息),我们给 3 只松鼠猴注射了 AZD8529(1 mg/kg,肌注),3 小时后,在轻度氯胺酮(10 mg/kg,肌注)麻醉下,从股静脉采集静脉血样本(约 1.5 ml)。我们迅速混合血样,并立即置于冰上冷却,直至离心。采血后30分钟内,将血液样本在4°C下以1500 xg离心10分钟制备血浆。分离血浆后,将其转移至两个2毫升的微量离心管中。血浆样本储存于-80°C。我们将样本用干冰运送至阿斯利康公司,该公司采用标准化的LC/MS/MS方法测定AZD8529的浓度。 大鼠体内微透析[1] 一般操作流程已在之前文献中描述(36)。在Sprague-Dawley大鼠中,于植入探针后20-24小时进行微透析,探针靶向伏隔核壳部(距前囟前2.0毫米,侧方1.1毫米,硬脑膜下8.0毫米)(37)。我们每隔20分钟采集一次样品(20 μl)(灌注速率:1 μl/min),并立即采用高效液相色谱-电化学检测法分析多巴胺水平。在连续3个样品中观察到多巴胺水平稳定(变化<15%)后,我们注射测试药物或其溶剂。在注射溶剂或尼古丁(0.4 mg/kg,皮下注射)前2小时,我们注射溶剂或AZD8529(10或30 mg/kg,腹腔注射)。尼古丁注射后,我们收集透析液样品2小时。我们根据阿斯利康之前在鼠行为模型中未发表的研究以及最近一项关于该药物对大鼠甲基苯丙胺渴求“潜伏期”影响的研究,确定了AZD8529的剂量。[1] AZD8529游离碱溶于无菌水中。[1] 实验设计[2] 实验1:急性AZD8418和AZD8529治疗对尼古丁和食物自我给药的影响[2] 在建立稳定的尼古丁或食物自我给药(连续三天反应变异性<20%)后,研究了急性AZD8418(0、0.37、1.12、3.73、7.46和)对尼古丁和食物自我给药的影响。采用受试者内拉丁方设计,评估了 AZD8418(14.92 mg/kg)和 AZD8529(0、1.75、5.83、17.5 和 58.3 mg/kg)治疗对尼古丁和食物自我给药的影响。使用四组未经处理的大鼠,分别考察了急性治疗对以下两组大鼠的影响:(i) AZD8418 对尼古丁自我给药(n = 12)和食物自我给药(n = 10)的影响;(ii) AZD8529 对尼古丁自我给药(n = 12)和食物自我给药(n = 7)的影响。两次给药之间至少间隔 5 天。 实验 2:长期 AZD8418 和 AZD8529 治疗对尼古丁和食物自我给药的影响 [2] 采用组间设计评估了 14 天重复 AZD8418 和 AZD8529 治疗方案对尼古丁和食物自我给药的影响。使用四组未经处理的大鼠来检验以下慢性治疗对大鼠的影响:(i)AZD8418(0、3.73、7.46 和 14.92 mg/kg/天)对尼古丁自我给药的影响(每组 n = 10–11);(ii)AZD8418(0 和 14.92 mg/kg/天)对食物自我给药的影响(每组 n = 10–11);(iii)AZD8529(0 和 58.3 mg/kg/天)对尼古丁自我给药的影响(每组 n = 10–12);以及(iv)AZD8529(0 和 58.3 mg/kg/天)对食物自我给药的影响(每组 n = 8–13)。在开始慢性治疗前,每个受试化合物的亚组均已根据体重和尼古丁/食物摄入量进行平衡。 实验 3:急性 AZD8418 和 AZD8529 治疗对线索诱导的尼古丁和食物渴求行为恢复的影响 [2] 完成实验 1 后,对尼古丁自我给药的大鼠进行消退测试。所有大鼠在第 10 次消退训练结束时均达到了预定的消退标准(见上文)。在给予载体后进行第一次恢复训练,以确保每个受试者在开始药物治疗前均表现出如上所述的显著恢复(与最后三次消退训练的平均值相比,反应增加 >50%)。只有在首次复吸实验中表现出强烈尼古丁渴求行为的大鼠才被纳入后续实验。每次复吸实验前均进行三天的消退实验,以再次消除大鼠的尼古丁渴求行为。在每次恢复实验前,分别给予AZD8418(0、1.12、3.73、7.46和14.92 mg/kg)和AZD8529(0、1.75、5.83、17.5和58.3 mg/kg),采用组内拉丁方设计。使用训练有素的独立大鼠(每组n = 9–13),这些大鼠已接受过食物自我给药训练,用于评估AZD8418(0、3.73、7.46和14.92 mg/kg)和AZD8529(0、1.75、5.83、17.5和58.3 mg/kg)对线索诱导的食物寻求行为恢复的影响。由于食物寻求行为在重复刺激后会迅速消退,因此采用组间设计。恢复性测试(Bespalov 等,2005)。治疗前,各组的体重和食物反应均保持平衡。 药代动力学研究[2] 将AZD8529(4.7 mg/kg)和AZD8418(5 mg/kg)通过灌胃法给予雄性Wistar大鼠(n = 3–4)组,单次给药或连续7天每日给药。在给药后第1天或第7天,分别于给药后0.5、1、3、6、12和24小时采集所有大鼠的尾静脉血样(0.25 ml)。血液样本采集后30分钟内,将血浆在4℃下以1500×g离心10分钟制备,并采用标准反相液相色谱-电喷雾电离串联质谱法(LC/MS/MS)分析AZD8529或AZD8418的浓度。 松鼠猴尼古丁自我给药:使用有尼古丁自我给药史的成年雄性松鼠猴。实验持续1小时,每周5天。在固定比率10(FR10,超时60秒)的强化程序下,按压10次操作杆后,将进行一次静脉注射尼古丁(30 µg/kg/次),并伴有2秒的视觉提示。在稳定的基线反应后,测试了AZD8529(0.03、0.3、1、3 和 10 mg/kg)或载体的效果。AZD8529在实验前 3 小时进行肌注。每个剂量连续测试 3 次,前后各 3 次均进行载体对照实验。[1] 松鼠猴食物自我给药:另一组猴子在相同的 FR10、60 秒暂停强化程序条件下自我给药 190 mg 食物颗粒。AZD8529(3、10 和 30 mg/kg,肌注)的测试采用与尼古丁实验相同的预处理和实验方案。[1] 松鼠猴恢复测试:在尼古丁强化反应消退后,进行恢复测试。对于尼古丁启动诱导的复吸实验,猴子在非条件性尼古丁(0.1 mg/kg,静脉注射)或生理盐水启动注射前3小时,接受AZD8529(0.3、1或3 mg/kg,肌肉注射)或载体。测试期间,按压操作杆(FR10)可获得生理盐水和线索。对于线索诱导的复吸实验,在尼古丁和线索均不可用的消退期后,猴子预先接受AZD8529或载体处理,然后按压操作杆(FR10)可获得先前与尼古丁相关的线索和生理盐水输注。[1] 大鼠微透析:将微透析探针植入雄性Sprague-Dawley大鼠的伏隔核壳部。恢复后,每20分钟收集一次透析液样本。建立稳定的基线多巴胺水平后,大鼠预先接受AZD8529(10或30 mg/kg,腹腔注射)或载体处理。两小时后,大鼠接受尼古丁(0.4 mg/kg,皮下注射)或载体注射。尼古丁注射后持续收集透析液2小时,并采用高效液相色谱-电化学检测法(HPLC-ECD)分析多巴胺水平。[1] 灵长类动物脑片放射自显影:食蟹猴麻醉后进行灌注,取出脑组织并冷冻。切取纹状体和海马的脑组织切片(20 µm)。将切片在检测缓冲液中预孵育,然后在含有 GDP、[35S]GTPγS 以及 LY379268(激动剂)、AZD8529(调节剂)和 LY341495(拮抗剂)不同组合的几种条件下孵育 2 小时。洗涤和干燥后,将切片曝光于胶片,显影并进行分析。[1] |
| 药代性质 (ADME/PK) |
AZD8529 的药代动力学特征 [2] AZD8418 单次给药 5 mg/kg 后,血浆峰浓度 (Tmax) 在给药后 1 小时达到。AZD8529 单次给药 4.7 mg/kg 后,血浆峰浓度在给药后 3 小时达到。AZD8418 的血浆峰浓度 (Cmax) (690 ± 108 nM) 显著高于 AZD8529 (158 ± 30 nM)。同样,AZD8418 的曲线下面积 (AUC) 也高于 AZD8529,表明 AZD8418 的生物利用度高于 AZD8529。连续 7 天每日重复给药 AZD8418 或 AZD8529 并未改变血浆暴露的 Tmax 或 Cmax。根据药代动力学研究结果,确定了AZD8418(0、0.37、1.12、3.73、7.46和14.92 mg/kg;预处理1小时)和AZD8529(0、1.75、5.83、17.5和58.3 mg/kg;预处理3小时)的给药剂量和预处理时间,以反映Cmax和Tmax的差异。
AZD8529的血浆浓度[2] 在一组松鼠猴(n=3)中,药物(1 mg/kg)注射后3小时(自我给药和恢复实验中的预处理时间),AZD8529的血浆浓度为112±17 nM。我们研究了全身注射 AZD8529 对尼古丁诱导的大鼠伏隔核壳部多巴胺释放的影响。结果显示,尼古丁(0.4 mg/kg,皮下注射)可增加细胞外多巴胺水平,而 30 mg/kg 的 AZD8529 可降低这种作用,但 10 mg/kg 的 AZD8529 则无此作用(补充图 S1;AZD8529 剂量与时间的交互作用:F(34,170)=2.24;p<0.001)。单独使用 AZD8529(10 或 30 mg/kg)对多巴胺水平无影响(补充图 S1)。在松鼠猴中,肌注 1 mg/kg AZD8529 3 小时后,血浆浓度为 112 ± 17 nM。该时间点与行为学实验中使用的预处理时间点一致。[1] 根据文中引用的公司信息,AZD8529 口服生物利用度高,且在人体研究中,其血浆浓度与猴子服用有效行为剂量后达到的浓度相当,耐受性良好。[1] |
| 毒性/毒理 (Toxicokinetics/TK) |
该手稿并未提供具体的毒性数据,例如半数致死量(LD50)、器官毒性或详细的药物相互作用。[1]
在高剂量(10 和 30 mg/kg)下,猴子的食物自主摄取量减少,其影响可能反映了行为抑制或其他副作用,但在本研究中,这些并未被归类为典型的毒性。[1] |
| 参考文献 |
|
| 其他信息 |
AZD8529 正在临床试验 NCT00921804(评估 AZD8529 在成人精神分裂症患者中的疗效、安全性和耐受性的研究)中进行研究。
背景:基于啮齿动物研究,II 型代谢型谷氨酸受体(mGluR2 和 mGluR3)被认为是成瘾治疗的靶点。然而,LY379268 和其他 II 型激动剂无法区分主要位于突触前抑制的 mGluR2(拟议的治疗靶点)和 mGluR3。这些激动剂在重复给药后会产生耐受性,因此不再被考虑用于成瘾治疗。本研究旨在探讨选择性mGluR2正向变构调节剂AZD8529对松鼠猴和大鼠尼古丁滥用相关效应的影响。方法:首先,我们利用表达人mGluR2的细胞系制备的膜以及灵长类动物脑片,通过体外功能性实验评估AZD8529对mGluR2功能的调节作用。然后,我们测定了AZD8529(0.03-10 mg/kg,肌注)对静脉注射尼古丁自我给药以及尼古丁启动或尼古丁相关线索诱导的尼古丁渴求恢复的影响。我们还测定了AZD8529对猴子食物自我给药行为的影响,以及对大鼠伏隔核壳部尼古丁诱导多巴胺释放的影响。[1] 原理:许多药物研发策略旨在通过阻断谷氨酸传递来减少尼古丁摄入量并防止吸烟复发。通过激活突触前抑制性代谢型谷氨酸(mGlu)2/3受体来减少谷氨酸释放,可以抑制尼古丁的强化作用,并阻断大鼠因线索诱导而恢复的尼古丁渴求行为。然而,mGlu2受体在尼古丁依赖中的相对作用仍不清楚。目的:本研究使用新型的、相对选择性的mGlu2正向变构调节剂(PAMs)AZD8418和AZD8529,评估mGlu2受体在尼古丁摄入和觅药行为中的作用。结果:AZD8418(0.37、1.12、3.73、7.46和14.92 mg/kg)和AZD8529(1.75、5.83、17.5和58.3 mg/kg)的急性给药可降低尼古丁的自我给药行为,但对食物维持的反应无影响。AZD8418的长期给药可减弱尼古丁的自我给药行为,但这种作用很快产生耐受性。长期服用AZD8529对尼古丁自我给药的抑制作用持续了14天的治疗。长期服用任何一种PAMs均可抑制食物自我给药。AZD8418(急性)和AZD8529(急性及亚慢性)均可阻断线索诱导的尼古丁和食物渴求行为的恢复。结论:这些发现表明mGlu2受体在自我给药尼古丁的强化作用以及先前与尼古丁给药相关的线索的动机影响(即线索诱导的尼古丁渴求行为的恢复)中发挥着重要作用。因此,mGlu2 PAMs可能有助于人们戒烟并预防复吸。[2] AZD8529被提出作为一种治疗尼古丁成瘾的新型疗法,其重点是通过靶向mGluR2来预防复吸。 [1] 与早期非选择性mGluR2/3正位激动剂(例如LY379268)相比,该药物具有潜在优势,因为它对mGluR2具有选择性,在测试模型中重复给药不会产生耐受性,并且在抗尼古丁剂量下对自然奖赏(食物)的影响极小。[1] 其作用机制被认为涉及减少中脑边缘通路中尼古丁诱导的多巴胺释放(如在大鼠中所示),并减弱尼古丁本身以及与尼古丁使用相关的环境线索的动机效应。[1] 鉴于共病性和共同的神经生物学通路,作者认为mGluR2正向变构调节剂的疗效可能不仅限于尼古丁成瘾,还可能扩展到其他物质使用障碍(例如可卡因、甲基苯丙胺、海洛因、酒精)。[1] |
| 分子式 |
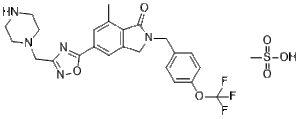
C25H28F3N5O6S
|
|---|---|
| 分子量 |
583.579935073853
|
| 精确质量 |
583.171
|
| 元素分析 |
C, 51.45; H, 4.84; F, 9.77; N, 12.00; O, 16.45; S, 5.49
|
| CAS号 |
1314217-69-0
|
| 相关CAS号 |
AZD-8529; 1092453-15-0; 1314217-69-0 (mesylate); 1344999-56-9 (mesylate hydrate)
|
| PubChem CID |
53495155
|
| 外观&性状 |
White to off-white solid powder
|
| tPSA |
147
|
| 氢键供体(HBD)数目 |
2
|
| 氢键受体(HBA)数目 |
13
|
| 可旋转键数目(RBC) |
6
|
| 重原子数目 |
40
|
| 分子复杂度/Complexity |
822
|
| 定义原子立体中心数目 |
0
|
| SMILES |
S(C)(=O)(=O)O.FC(OC1C=CC(=CC=1)CN1C(C2C(C)=CC(C3=NC(CN4CCNCC4)=NO3)=CC=2C1)=O)(F)F
|
| InChi Key |
HRHCPVNKPOTCGB-UHFFFAOYSA-N
|
| InChi Code |
InChI=1S/C24H24F3N5O3.CH4O3S/c1-15-10-17(22-29-20(30-35-22)14-31-8-6-28-7-9-31)11-18-13-32(23(33)21(15)18)12-16-2-4-19(5-3-16)34-24(25,26)27;1-5(2,3)4/h2-5,10-11,28H,6-9,12-14H2,1H3;1H3,(H,2,3,4)
|
| 化学名 |
methanesulfonic acid;7-methyl-5-[3-(piperazin-1-ylmethyl)-1,2,4-oxadiazol-5-yl]-2-[[4-(trifluoromethoxy)phenyl]methyl]-3H-isoindol-1-one
|
| 别名 |
AZD-8529; AZD8529 mesylate; AZD 8529 mesylate; 1314217-69-0; AZD-8529 (mesylate); 29K1DH0MV7; 7-Methyl-5-(3-(piperazin-1-ylmethyl)-1,2,4-oxadiazol-5-yl)-2-(4-(trifluoromethoxy)benzyl)isoindolin-1-one methanesulfonate; methanesulfonic acid;7-methyl-5-[3-(piperazin-1-ylmethyl)-1,2,4-oxadiazol-5-yl]-2-[[4-(trifluoromethoxy)phenyl]methyl]-3H-isoindol-1-one; 1H-Isoindol-1-one, 2,3-dihydro-7-methyl-5-(3-(1-piperazinylmethyl)-1,2,4-oxadiazol-5-yl)-2-((4-(trifluoromethoxy)phenyl)methyl)-, methanesulfonate (1:1); AZD-8529 mesylate
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month 注意: 请将本产品存放在密封且受保护的环境中,避免吸湿/受潮。 |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO: ~41.7 mg/mL (~71.4 mM)
|
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.08 mg/mL (3.56 mM) (饱和度未知) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 20.8 mg/mL澄清DMSO储备液加入400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: ≥ 2.08 mg/mL (3.56 mM) (饱和度未知) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 例如,若需制备1 mL的工作液,可将 100 μL 20.8 mg/mL澄清DMSO储备液加入900 μL 20% SBE-β-CD生理盐水溶液中,混匀。 *20% SBE-β-CD 生理盐水溶液的制备(4°C,1 周):将 2 g SBE-β-CD 溶解于 10 mL 生理盐水中,得到澄清溶液。 View More
配方 3 中的溶解度: ≥ 2.08 mg/mL (3.56 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 1.7136 mL | 8.5678 mL | 17.1356 mL | |
| 5 mM | 0.3427 mL | 1.7136 mL | 3.4271 mL | |
| 10 mM | 0.1714 mL | 0.8568 mL | 1.7136 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
| NCT Number | Recruitment | interventions | Conditions | Sponsor/Collaborators | Start Date | Phases |
| NCT00755378 | Completed | Drug: AZD8529 Drug: Placebo |
Healthy Volunteer | AstraZeneca | September 2008 | Phase 1 |
| NCT00765492 | Completed | Drug: AZD8529 Drug: Placebo |
Healthy Volunteer | AstraZeneca | October 2008 | Phase 1 |
| NCT00886756 | Completed | Drug: AZD8529 Drug: Placebo |
Healthy | AstraZeneca | April 2009 | Phase 1 |
| NCT02401022 | Completed | Drug: AZD8529 | Tobacco Use Disorder | National Institute on Drug Abuse (NIDA) |
July 2015 | Phase 2 |
| NCT00921804 | Completed | Drug: AZD 8529 Drug: Risperidone |
Schizophrenia | AstraZeneca | June 2009 | Phase 2 |
|
AZD8529 decreased nicotine priming-induced and cue-induced reinstatement in squirrel monkeys.Biol Psychiatry. 2015 Oct 1;78(7):452-62. |
Effect of AZD8529 on nicotine and food self-administration in squirrel monkeys.Biol Psychiatry. 2015 Oct 1;78(7):452-62. |
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1
































 COA
COA



 463611831
463611831
