| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| Other Sizes |
|

| 靶点 |
Piezo1
|
|---|---|
| 体外研究 (In Vitro) |
在 HEK 293 和 CHO 细胞中,Dooku1(10 μM,300 秒)表现出对 Piezo1 通道的选择性 [1]。在 Piezo1 T-REx 细胞中,Dooku1(10 μM,140 秒)对 Piezo1 通道的组成型活性没有影响 [1]。在 HEK 293 和 Piezo1 T-REx 细胞中,Dooku1(10 μM,40-60 秒)会阻断内源性 Yoda1 激活通道 [1]。
Yoda1吡嗪环的修饰产生了一种类似物,该类似物缺乏激动剂活性,但可逆地拮抗Yoda1。这种类似物被称为Dooku1。Dooku1抑制了2μM Yoda1诱导的Ca2+进入,IC50为1.3μM(HEK 293细胞)和1.5μM(HUVEC),但未能抑制组成型Piezo1通道活性。它对内源性ATP诱发的HEK 293细胞中Ca2+升高或储存操作的Ca2+进入,或通过CHO和HEK 293电池中过表达的TRPV4或TRPC4通道进入Ca2+没有影响[1]。 |
| 体内研究 (In Vivo) |
Dooku1(10 μM 动脉松弛 20 分钟)阻断可降低 Yoda1 诱导的野生型 C57BL/6 小鼠的主动脉松弛 [1]。
Dooku1抑制Yoda1诱导的主动脉舒张[1] 为了确定Dooku1是否抑制Yoda1引起的舒张,将主动脉环与10μM Dooku1预孵育20分钟。Dooku1强烈抑制了Yoda1诱导的舒张(图8A-C)。为了更详细地描述这一现象,我们在主动脉测定中测试了另外四种Yoda1类似物。所选的类似物显示出抑制Piezo1-T-REx细胞中Yoda1反应的各种能力:类似物2e(无激活和无抑制)(图1)、2g(轻微激活和轻微抑制)(见图1)、7b(轻微活化和部分抑制)(附图2和3)和11。模拟2e没有影响(图8D-F)。相比之下,2g、7b和11抑制了Yoda1诱导的弛豫(图8G-K)。此外,这些类似物抑制Yoda1诱导的弛豫的能力与抑制Yoda 1诱导的Ca2+进入相关(图8L)。数据表明,Dooku1作为Yoda1诱导的主动脉舒张抑制剂具有很强的疗效,该舒张是通过破坏Yoda1诱发的Piezo1通道活性介导的。 Dooku1对Yoda1诱导的舒张具有选择性,但部分抑制激动剂收缩反应[1] 在Dooku1存在的情况下对PE反应的分析显示,对基线张力没有影响,但有显著的抑制作用(图9A,B)。为了确定Dooku1对PE诱导的收缩的抑制是否是这种收缩剂特有的,我们还测试了Dooku1对抗Tx A2模拟物U46619诱导的收缩作用。主动脉环用0.1μM U46619预收缩(图9C,D)。添加Dooku1导致部分松弛(图9D,E)。相比之下,Dooku1对ACh(1μM)或no供体SIN-1(10μM)诱发的舒张没有影响(图9F,G)。在其他四种Yoda1类似物存在的情况下,对PE反应的研究表明没有抑制作用(图10)。数据表明,Dooku1选择性抑制Yoda1诱导的舒张,但也通过未知机制部分抑制受体介导的激动剂反应。 |
| 酶活实验 |
细胞内Ca2+测量[1]
实验前24小时,将HEK 293和CHO细胞以90%的融合率铺在聚赖氨酸涂层96孔板(美国纽约州康宁)中,将HUVEC铺在透明96孔板中。在标准浴溶液(SBS)中的0.01%普朗尼克酸存在下,将细胞与2μM fura-2-AM或4μM fluo-4-AM(用于表达TRPV4的CHO细胞)在37°C下孵育1小时。对于氟-4的记录,在整个实验过程中,SBS中包括2.5mM丙磺舒。在室温下用SBS洗涤细胞30分钟。如果正在测试抑制剂,则应在SBS洗涤后立即添加这些抑制剂,并在实验的其余部分进行维护。在室温下,在Softmax Pro软件v5.4.5控制的96孔荧光板阅读器上进行测量。对于使用fura-2的记录,细胞内钙的变化(Δ)表示为340和380 nm激发下fura-2发射(510 nm)强度的比值。对于使用fluo-4的记录,染料在485nm处被激发,在525nm处发出收集的光,测量结果以任意单位显示为绝对荧光。SBS含有(mM):130 NaCl、5 KCl、8 D-葡萄糖、10 HEPES、1.2 MgCl2、1.5 CaCl2,用NaOH滴定pH至7.4。对于Ca2+加反实验,使用无Ca2+的SBS(不含CaCl2),Ca2+加回为0.3 mM。对于洗脱实验,在记录前立即用SBS洗涤抑制剂3次。 FluxOR™细胞内Tl+(铊离子)测量[1] 实验前24小时,将诱导(Tet+)和非诱导(Tet-)Piezo1 HEK 293细胞以90%的融合率铺在聚赖氨酸涂层的96孔板上,将HUVEC铺在透明96孔板中。细胞在室温下用FluxOR染料加载1小时,然后转移到测定缓冲液中20分钟。如果正在测试抑制剂,则此时添加这些抑制剂并在整个实验过程中保持。根据制造商的说明,用含Tl+的无K+溶液刺激细胞。在室温下,在Softmax Pro软件v5.4.5控制的96孔荧光板阅读器上进行测量。FluxOR在485nm处激发,在520nm处收集发射光,测量值表示为比基线增加的比率(F/F0)。 |
| 细胞实验 |
细胞活力测定 [1]
细胞类型: HUVEC、Piezo1 T-REx 细胞 测试浓度: 10 μM 孵育时间: 40-60 s 实验结果: 对Yoda1诱导的Ca2+进入HUVEC具有浓度依赖性抑制作用,IC50为1.49 μM。在 HUVEC 中的效力增强,EC50 为 0.23 μM,在 Piezo1 T-REx 细胞中的效力增强,EC50 为 2.51 μM。 |
| 动物实验 |
动物/疾病模型:野生型雄性C57BL/6小鼠主动脉环[1]
剂量:10 μM 给药途径:20分钟 实验结果:抑制Yoda1诱导的舒张。 动物[1] 实验采用12至16周龄的野生型雄性C57BL/6小鼠。所有小鼠均饲养于GM500独立通风笼中,温度为21°C,湿度为50-70%,光照/黑暗周期为12小时交替。小鼠可自由摄取RM1饲料,垫料为Pure'o Cell公司的产品。所有动物实验均已获得利兹大学动物伦理委员会和英国政府内政部的批准。动物实验报告遵循 ARRIVE 指南(Kilkenny 等,2010;McGrath 和 Lilley,2015)。 主动脉收缩研究[1] 使用小鼠血管的线张力测定技术被认为是研究血管反应性的有效模型(Outzen 等,2015)。解剖出胸主动脉后,立即将其置于冰冷的 Krebs 溶液(125 mM NaCl、3.8 mM KCl、1.2 mM CaCl₂、25 mM NaHCO₃、1.2 mM KH₂PO₄、1.5 mM MgSO₄、0.02 mM EDTA 和 8 mM D-葡萄糖,pH 7.4)中。在解剖显微镜下小心去除结缔组织和脂肪。将1毫米长的血管段安装在等长线张力测定系统中,该系统配备两根直径为40微米的钢丝,浸泡在37℃的克氏液中,并通入95%氧气和5%二氧化碳的混合气体。然后,将血管段逐步拉伸至其最佳静息张力,达到90%等效跨壁压(100 mmHg),并在实验前平衡1小时。该拉伸程度与舒张压时的预期拉伸程度大致相同(Rode等人,2017)。 |
| 参考文献 | |
| 其他信息 |
背景与目的:机械敏感性Piezo1通道在血管生理和疾病中发挥着重要作用。Yoda1是一种小分子激动剂,但除此之外,人们对这些通道的药理学研究还很有限。实验方法:采用合成化学方法制备了Yoda1类似物。在过表达通道亚基的HEK 293或CHO细胞系以及天然表达Piezo1的HUVEC细胞中测量了细胞内Ca2+和Tl+浓度。从小鼠胸主动脉环中记录了等长张力。主要结果:修饰Yoda1的吡嗪环得到了一种类似物,该类似物缺乏激动剂活性,但可逆地拮抗Yoda1。该类似物被命名为Dooku1。 Dooku1抑制了2 μM Yoda1诱导的Ca2+内流,IC50值分别为1.3 μM(HEK 293细胞)和1.5 μM(HUVECs细胞),但未能抑制组成型Piezo1通道活性。它对HEK 293细胞中内源性ATP诱导的Ca2+升高或储存操纵性Ca2+内流,以及在CHO和HEK 293细胞中过表达的TRPV4或TRPC4通道介导的Ca2+内流均无影响。 Yoda1 引起主动脉环剂量依赖性舒张,该作用由内皮依赖性和 NO 依赖性机制介导,并可被 Dooku1 及其类似物拮抗。结论和意义:Yoda1 诱导的 Piezo1 通道活性可能存在化学拮抗作用,提示存在具有不同结合域和效力域的特定化学相互作用位点。[1]
目前,仅有的 Piezo1 活性抑制剂对 Piezo1 不具有选择性(Drew 等,2002;Bae 等,2011)。Dooku1 也并非完美,因为它不能直接阻断通道,但它是一种可用于 Piezo1 表征研究的新型工具化合物。它拮抗 Yoda1 的作用,并可能有助于理解 Piezo1 通道上或附近重要的小分子结合位点。 Dooku1 在不具备激动剂活性的情况下,能有效抑制 Yoda1 诱导的 Piezo1 活性。它抑制 Piezo1 活性的同时,并不干扰细胞内其他 Ca2+ 处理过程,也不影响其他主动脉舒张剂的作用。尽管这些数据表明 Dooku1 对 Piezo1 通道具有特异性,但仍需进一步研究来验证这一点,尤其考虑到 Dooku1 对 PE 和 U46619 诱导的主动脉环收缩具有抑制作用,这可能反映了 Dooku1 的 Piezo1 机制或其他未知作用。Dooku1 可能作用于血管平滑肌细胞中的 Piezo1,部分抑制收缩。这假设在收缩过程中,这些通道通过类似 Yoda1 的机制被激活。研究发现,Piezo1 并非维持正常肌源性张力所必需(Retailleau 等,2015),因此,应考虑 Dooku1 是否存在非 Piezo1 靶点。 [1] Dooku1 仅对 Yoda1 诱导的 Piezo1 通道活性有效,而对组成型 Piezo1 通道活性无效。这种效应与 Dooku1 作用于与 Yoda1 相同或相似的位点,从而阻断 Yoda1 与其激动剂结合位点的结合相一致。Dooku1 的可逆性与 Yoda1 的可逆性一致(Rocio Servin-Vences 等,2017)。研究 Dooku1 的作用是否符合竞争性拮抗作用是有益的,但由于化合物的溶解度限制,无法构建合适的浓度-效应曲线。Dooku1 对组成型活性无任何影响,表明其背景通道活性的机制与 Yoda1 化学激活的机制不同。[1] Dooku1 在 HUVEC 细胞中部分抑制 Yoda1,但在主动脉中强烈抑制 Yoda1(图 6D 对比图 8C)。我们最初推测这种差异是由于收缩实验的温度较高(37°C 对比室温)造成的,但 Dooku1 效应与温度的依赖性并不显著(图 3K)。另一种解释可能是 Ca2+ 内流与 NO 生成并非直接成正比,因此部分抑制 Yoda-1 诱导的 Ca2+ 内流足以抑制 Yoda1 诱导的大部分舒张作用。另一个差异是,Yoda1 在 HUVEC 细胞中的效力高于 Piezo1 T-REx 细胞,这表明天然 Piezo1 通道和过表达 Piezo1 通道之间存在差异(图 6E、F)。我们推测,这种差异反映了内皮细胞中通道更高的基础活性状态,如前所述(Rode等人,2017),使得通道对Yoda1更敏感,因为它们更容易被激活。[1] 总之,本研究为Yoda1的构效关系提供了重要的见解,并支持了Piezo1通道上或其附近存在特定化学结合位点的观点。此外,本研究还发现了一种有用的工具化合物Dooku1,它能有效拮抗Yoda1诱导的Piezo1通道活性,从而将其与组成型Piezo1通道活性区分开来。 Piezo1在血管生物学中的完整作用仍在研究中,但该蛋白可能具有重要的临床意义,其在遗传疾病、血压控制、高血压诱导的动脉重塑和运动能力等方面发挥着重要作用(Retailleau et al., 2015; Wang et al., 2016; Rode et al., 2017)。目前尚不清楚激活或抑制该通道是否更有利,但如果未来要充分发挥该蛋白的治疗潜力,就必须加深我们对Piezo1的药理学和生理学认识。深入了解Piezo1通道与小分子的相互作用,有望成为全面理解Piezo1生物学的重要方面。[1] |
| 分子式 |
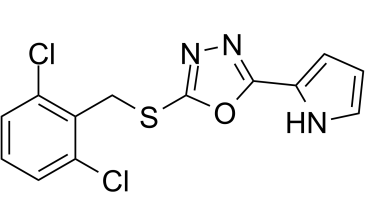
C13H9CL2N3OS
|
|---|---|
| 分子量 |
326.201059103012
|
| 精确质量 |
324.984
|
| CAS号 |
2253744-54-4
|
| PubChem CID |
137321150
|
| 外观&性状 |
White to off-white solid powder
|
| LogP |
3.8
|
| tPSA |
80
|
| 氢键供体(HBD)数目 |
1
|
| 氢键受体(HBA)数目 |
4
|
| 可旋转键数目(RBC) |
4
|
| 重原子数目 |
20
|
| 分子复杂度/Complexity |
316
|
| 定义原子立体中心数目 |
0
|
| SMILES |
ClC1C=CC=C(C=1CSC1=NN=C(C2=CC=CN2)O1)Cl
|
| InChi Key |
MNPOBXLPCWFONX-UHFFFAOYSA-N
|
| InChi Code |
InChI=1S/C13H9Cl2N3OS/c14-9-3-1-4-10(15)8(9)7-20-13-18-17-12(19-13)11-5-2-6-16-11/h1-6,16H,7H2
|
| 化学名 |
2-[(2,6-dichlorophenyl)methylsulfanyl]-5-(1H-pyrrol-2-yl)-1,3,4-oxadiazole
|
| 别名 |
Dooku1; Dooku-1; Dooku 1; 2253744-54-4; 2-((2,6-Dichlorobenzyl)thio)-5-(1H-pyrrol-2-yl)-1,3,4-oxadiazole; 2-[(2,6-Dichlorobenzyl)thio)-5-(1H-pyrrol-2-yl)-1,3,4-oxadiazole; 2-{[(2,6-dichlorophenyl)methyl]sulfanyl}-5-(1H-pyrrol-2-yl)-1,3,4-oxadiazole; 2-((2,6-Dichlorobenzyl)thio)-5-(1H-pyrrol-2-yl)-1,3,4- oxadiazole; Dooku 1
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month 注意: 请将本产品存放在密封且受保护的环境中,避免吸湿/受潮。 |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO : ~100 mg/mL (~306.56 mM)
|
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.08 mg/mL (6.38 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 20.8 mg/mL 澄清 DMSO 储备液加入到 900 μL 玉米油中并混合均匀。 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 3.0656 mL | 15.3280 mL | 30.6560 mL | |
| 5 mM | 0.6131 mL | 3.0656 mL | 6.1312 mL | |
| 10 mM | 0.3066 mL | 1.5328 mL | 3.0656 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
 Piezo1 agonist 1-d2
Piezo1 agonist 1-d2
 Yaddle1
Yaddle1
 D-GsMTx4
D-GsMTx4
 D-GsMTx4 TFA
D-GsMTx4 TFA
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 463611831
463611831
