| 规格 | 价格 | |
|---|---|---|
| 500mg | ||
| 1g | ||
| Other Sizes |

| 靶点 |
S1P2 (Sphingosine-1-Phosphate 2; EDG-5); human S1P2 (IC50 = 17 nM); rat S1P2 (IC50 = 22 nM)
|
||
|---|---|---|---|
| 体外研究 (In Vitro) |
体外活性:JTE-013 逆转 S1P2 信号传导对血管 EC 和平滑肌细胞细胞迁移的抑制作用。它还在体外调节内皮紧密连接和屏障功能。 JTE-013 阻断 S1P2 信号传导可显着增强 S1P 对 TEER(内皮完整性的体外测量)增加以及衰老 EC 中 TJ 形成的影响 激酶测定:JTE 013 是一种新型有效且选择性的 S1P2 (sphingosine-1-磷酸 2) 拮抗剂,IC50 为 17.6 nM。它与人和大鼠受体结合的 IC50 值分别为 17 和 22 nM,对人 S1P1 和 S1P3 的 IC50 值 >10 µM。它逆转 S1P2 信号传导对血管 EC 和平滑肌细胞细胞迁移的抑制作用。它还在体外调节内皮紧密连接和屏障功能。 JTE-013 阻断 S1P2 信号传导可显着增强 S1P 对 TEER(内皮完整性的体外测量)增加以及衰老 EC 中 TJ 形成的影响。细胞测定:与不同浓度的 JTE-013 预孵育 10 分钟的 HUVEC 可以向放置指定浓度的 Sph-1-P 的下室迁移 4 小时。将过滤器下侧的迁移细胞固定、染色并计数。[2]
JTE-013(50-200 μM;1-3 天)会降低细胞活力[1]。 JTE-013(10-1000 nM;30 分钟)可逆转 S1P 诱导的 Akt 抑制并抑制 S1P 诱导的 ERK 激活[1]。 JTE-013 对 S1P3 具有 4.2% 的抑制作用,并且在浓度高达 10 μM 时不会拮抗 S1P1[1]。hr> 细胞活力测定[1] 细胞系:SK-N-AS 细胞 浓度:50、100、150、200 μM 孵育时间:1-3 天 结果:细胞活力降低。 Western Blot 分析[1] 细胞系:SK-N-AS 细胞 浓度:10、100、1000 nM 孵育时间:30 分钟 结果:逆转 S1P 诱导的 Akt 抑制并抑制 S1P 诱导的 ERK 激活。 JTE-013对GB细胞迁移的影响[1] S1P2信号转导的一个很好的生物学后果是抑制细胞迁移(Lepley等,2005)。为了比较JTE-013和AB1作为S1P2拮抗剂的效率,进行了细胞迁移实验。使用GB细胞系U118和U87,因为众所周知,S1P抑制S1P2高表达的U118细胞的细胞迁移,而S1P诱导S1P2低表达的U87细胞的细胞迁移(Lepley et al., 2005;图3,A和C)。在逆转U118细胞中s1p介导的细胞迁移抑制和通过阻断S1P2信号传导增强U87细胞中s1p刺激的细胞迁移方面,AB1处理比JTE-013更有效(图3,B和D)。 AB1与JTE-013对SK-N-AS细胞S1P2下游分子水平的影响[1] S1P2通过调控不同的下游效应分子发挥多种细胞功能。我们之前的研究以及其他人的研究表明,这些分子包括细胞内信号介质(p-Akt, p-ERK),以及生长和分化调节剂,如CTGF (Sanchez et al., 2007;Li et al., 2008a)。为了进一步比较JTE-013和AB1的体外效率,我们进行了Western blot分析。与JTE-013类似,AB1能够逆转s1p诱导的Akt抑制,并抑制s1p诱导的ERK激活,浓度在100 nM至1 μM之间(图4A)。实时定量聚合酶链反应进一步显示AB1比JTE-013在抑制s1p诱导的CTGF mRNA表达方面相对更有效(图4B)。 AB1与JTE-013对SK-N-AS细胞活力的影响[1] 为了研究AB1抑制肿瘤作用的潜在机制,我们对JTE-013或AB1处理的SK-N-AS细胞的细胞活力进行了评估。MTT实验显示,在SK-N-AS细胞中,浓度高于50 μM时,AB1对细胞活力的抑制作用不如JTE-013,而在较低浓度时,AB1的抑制作用相似(图6),这表明AB1引起的抑制效果的改善不是由直接抑制癌细胞的细胞存活引起的。 |
||
| 体内研究 (In Vivo) |
JTE-013 对 S1P2 的抑制可显着抑制体内动物模型中的微血管通透性。它通过抑制脑血管通透性、脑内出血的发展和中风实验模型中的神经血管损伤来调节脑内皮的反应。在体外和体内研究中,JTE-013 降低了肥大细胞活化、气道浸润以及组胺和多种细胞因子的血清水平。在小鼠模型中,JTE-013 抑制链脲佐菌素诱导的血糖升高、胰腺 b 细胞凋亡和糖尿病的发病率。在高脂肪饮食条件下的新西兰肥胖糖尿病小鼠模型中,它可以保护胰腺 b 细胞。 JTE-013 治疗还可降低 ApoE−/− 小鼠中 IL-1b 和 IL-18(内毒素诱导的炎症细胞因子)的血浆水平,并且 S1P2 基因缺陷可减少动脉粥样硬化。该化合物提供了一种治疗动脉粥样硬化和败血症等炎症性疾病的新方法。
JTE-013(灌胃;每天 30 mg/kg,连续 14 天)可减小肿瘤大小和肿瘤重量[1]。 动物模型:六周龄雌性无胸腺NCr-nu/nu裸鼠[1] 剂量:30 mg/kg 给药方式:灌胃;连续 14 天每天一次 结果:肿瘤大小和重量减小。 JTE-013的对比[1] 在之前的研究中,JTE-013被证明可以显著抑制NB异种移植物的生长(Li et al., 2011)。在这里,发现AB1在抑制NB异种移植物生长方面再次比JTE-013更有效,无论是肿瘤大小(图5A)还是治疗后14天的肿瘤重量(图5B)。综上所述,上述数据有力地表明,与JTE-013相比,AB1可能具有更强的体内抗肿瘤活性。 AB1与JTE-013对NB异种移植物CCL2表达和肿瘤相关巨噬细胞浸润的影响[1] 为了阐明AB1增强抗肿瘤作用的机制,研究人员量化了AB1对处理过的NB异种移植物中几种S1P2下游分子基因表达水平的影响。CCL2就是这些基因之一(Li et al., 2014)。CCL2的表达与肿瘤相关巨噬细胞(TAM)浸润呈正相关(Zhang et al., 2010)。在我们之前的研究中,JTE-013阻断S1P2信号不仅抑制NB异种移植物的生长(Li et al., 2011),而且还降低了CCL2的表达和随后的TAM浸润(Li et al., 2014),表明抑制CCL2有利于抗癌治疗。正如预期的那样,在JTE-013处理或AB1处理的NB异种移植物中,CCL2在mRNA和蛋白水平上的表达都有降低的趋势(图7,A和B)。此外,小鼠巨噬细胞标志物F4/80的免疫组织化学染色显示,JTE-013和AB1均显著抑制TAM的浸润。然而,AB1没有表现出任何改善的抑制作用(图7C),这表明AB1抗肿瘤作用的增强不是由于影响CCL2表达和随后的TAM浸润。 AB1与JTE-013对NB异种移植物肿瘤纤维化和凋亡的影响[1] CTGF是纤维化的中心介质(Lipson et al., 2012)。有趣的是,在mRNA和蛋白水平上,AB1比JTE-013抑制CTGF表达的程度更大(图8),这表明AB1抗肿瘤效果的提高可能部分是由于其对肿瘤纤维化的有益作用。使用Ki67染色的组织学研究未发现三组之间Ki67阳性增殖细胞的数量有显著差异(补充图2)。然而,末端脱氧核苷酸转移酶介导的地高西根-脱氧尿苷镍端标记染色(图9A)和cleaved caspase-3检测(图9B)显示,AB1在诱导这些NB异种移植物上的肿瘤细胞凋亡方面比JTE-013更有效。综上所述,我们的数据表明,AB1引起的抗肿瘤效果的提高可能归因于对NB异种移植物肿瘤纤维化和肿瘤凋亡的影响。 |
||
| 酶活实验 |
荧光成像平板阅读器试验。[1]
钙通量测定由一家CRO公司在FLIPRTETRA仪器上进行[荧光成像板阅读器(flir)测定],以分析测试化合物在S1P1-5上的剂量依赖性激动剂和拮抗剂活性。简单地说,激动剂实验是在FLIPRTETRA仪器上进行的,在荧光基线建立后,将测试化合物、对照物和参比激动剂S1P添加到检测板上。180秒的时间用来评估每种化合物激活每种S1PR的能力。激动剂实验完成后,从FLIPRTETRA仪器中取出检测板,在25°C下孵育7分钟。之后,将检测板放回FLIPRTETRA仪器中,开始拮抗剂检测。利用激动剂试验中确定的EC80效价值,在建立荧光基线后,用参比激动剂S1P的EC80浓度挑战所有预孵育的样品复合孔。另外180秒的时间用来评估每种化合物抑制每种S1PR的能力。所有分析板数据都进行了适当的基线校正。应用基线校正后,导出最大荧光值,并处理数据以计算激活百分比(相对于Emax参考激动剂S1P和载体控制值)和抑制百分比(相对于EC80和载体控制值)。所有剂量反应曲线均采用GraphPad Prism软件生成 |
||
| 细胞实验 |
细胞系:SK-N-AS 细胞 浓度:50、100、150、200 μM 孵育时间:1-3 天 结果:细胞活力降低。
细胞系:SK-N-AS细胞
浓度:50、100、150、200 μM 孵育时间:1-3天 结果:细胞活力降低, 迁移试验。[1] 如前所述,迁移试验在96孔趋化微室中进行(Li等人,2009b)。简单地说,聚碳酸酯过滤器(孔径为8µm)涂覆50µg/ml纤维连接蛋白。S1P稀释后以85µl /孔加入下腔。胰蛋白酶化前对GB细胞进行血清饥饿2小时,用或不加<强JTE-013和AB1预处理10分钟。在0.39 ml培养基中,每孔5 × 104个细胞置于上隔室,在37℃下迁移5小时。然后将过滤器在4°C下固定过夜,用棉签去除未迁移的细胞。在96孔板上用0.1%结晶紫染色,10%醋酸洗脱。在595 nm处测定吸光度。 甲基噻唑基二苯基溴化四氮唑测定[1] 用JTE-013或AB1处理的SK-N-AS细胞的活力通过甲基噻唑基二苯基溴化四唑(MTT)测定,如先前所述(Li et al., 2013)。简单地说,将SK-N-AS细胞接种于96孔板中,用不同浓度的JTE-013或AB1处理不同时间,然后在37℃下MTT孵育2小时。用二甲基亚砜溶解活细胞中形成的不溶性甲醛,用Bio-Rad微孔板仪在595 nm处测定吸光度。结果显示为细胞活力相对于非药物治疗对照的百分比。 |
||
| 动物实验 |
|
||
| 参考文献 | |||
| 其他信息 |
生物活性脂质鞘氨醇-1-磷酸(S1P)及其受体(S1P1-5)在包括癌症在内的多种病理过程中发挥着关键作用。S1P轴已成为癌症治疗的重要靶点。已知的S1P2拮抗剂JTE-013 [N-(2,6-二氯-4-吡啶基)-2-[1,3-二甲基-4-(1-甲基乙基)-1H-吡唑并[3,4-b]吡啶-6-基]-肼甲酰胺]在体内稳定性较差。因此,开发结构修饰、效力更强、稳定性更高的S1P2抑制剂将是理想的药理学工具。 JTE-013 的衍生物之一 AB1 [N-(1H-4-异丙基-1-烯丙基-3-甲基吡唑并[3,4-b]吡啶-6-基)-氨基-N'-(2,6-二氯吡啶-4-基)脲] 与 JTE-013 相比,表现出更强的 S1P2 拮抗活性。静脉注射药代动力学研究表明,AB1 在体内稳定性增强或清除速度减慢。胶质母细胞瘤细胞迁移实验表明,AB1 在阻断 S1P2 介导的细胞迁移抑制方面略优于 JTE-013。在神经母细胞瘤 (NB) 细胞系 SK-N-AS 中的功能研究表明,AB1 在影响 S1P2 下游信号分子方面至少与 JTE-013 具有同等效力。同样,与 JTE-013 相比,AB1 对 SK-N-AS 肿瘤异种移植瘤生长的抑制作用也更强。细胞活力检测排除了AB1增强作用是由抑制癌细胞存活所致的可能性。JTE-013和AB1均有抑制(CC基序)配体2表达的趋势,并能显著抑制NB异种移植瘤中后续的肿瘤相关巨噬细胞浸润。值得注意的是,AB1在抑制促纤维化介质结缔组织生长因子表达方面比JTE-013更有效。末端脱氧核苷酸转移酶介导的地高辛-脱氧尿苷缺口末端标记法(TUNEL)和裂解型caspase-3检测进一步证实,与JTE-013相比,AB1处理的NB异种移植瘤中细胞凋亡增加。总而言之,JTE-013的修饰产物AB1提高了其效力、静脉药代动力学、细胞活性和抗肿瘤活性,并可能增强其临床和实验应用价值。 [1]
总之,我们在此报告了S1P2拮抗剂JTE-013的新型修饰产物AB1。AB1的效力和静脉药代动力学特性均有适度提高,稳定性更佳。在神经母细胞瘤(NB)的背景下,AB1似乎也具有更好的细胞活性和抗肿瘤活性。基于这些发现,我们得出结论,AB1可能具有更强的临床和实验应用价值,克服了JTE-013的一些不足。[1] 鞘氨醇-1-磷酸(S1P)调节培养的内皮细胞中的多种分子和细胞事件,例如细胞骨架重塑、细胞-细胞外基质相互作用和细胞间连接相互作用。我们利用Sprague-Dawley大鼠提睾肌血管床的静脉渗漏模型,研究了S1P信号在微血管通透性调节中的作用。 S1P信号传导由S1P家族G蛋白偶联受体(S1P(1-5)受体)介导。S1P(1)和S1P(2)受体分别传递刺激性和抑制性信号,它们在提睾肌血管内皮细胞中表达。在Sprague-Dawley大鼠中,单独经颈动脉注射S1P无法保护提睾肌血管床免受组胺诱导的静脉渗漏。然而,使用S1P(1)的两种激动剂SEW2871和FTY720激活S1P(1)介导的信号传导,可显著抑制组胺诱导的微血管渗漏。使用VPC 23019拮抗S1P(1)调节的信号传导,则显著增强了组胺诱导的静脉渗漏。使用S1P(2)特异性拮抗剂JTE-013抑制S1P(2)信号通路后,S1P能够在体内保护微血管通透性。此外,在培养的内皮细胞中,内皮细胞紧密连接和屏障功能受S1P(1)和S1P(2)介导的信号通路协同调节。这些数据表明,S1P(1)和S1P(2)信号通路之间的平衡调节外周循环中微血管通透性的稳态,从而可能影响外周血管总阻力。[2] |
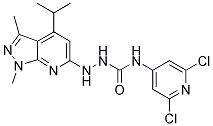
| 分子式 |
C17H19CL2N7O
|
|
|---|---|---|
| 分子量 |
408.29
|
|
| 精确质量 |
407.102
|
|
| 元素分析 |
C, 50.01; H, 4.69; Cl, 17.37; N, 24.01; O, 3.92
|
|
| CAS号 |
547756-93-4
|
|
| 相关CAS号 |
547756-93-4
|
|
| PubChem CID |
25168534
|
|
| 外观&性状 |
Typically exists as
White to off-white solid at room temperature
|
|
| 密度 |
1.5±0.1 g/cm3
|
|
| 折射率 |
1.697
|
|
| LogP |
4.42
|
|
| tPSA |
96.76
|
|
| 氢键供体(HBD)数目 |
3
|
|
| 氢键受体(HBA)数目 |
5
|
|
| 可旋转键数目(RBC) |
5
|
|
| 重原子数目 |
27
|
|
| 分子复杂度/Complexity |
504
|
|
| 定义原子立体中心数目 |
0
|
|
| SMILES |
CN1C2=NC(NNC(NC3C(Cl)=CN=CC=3Cl)=O)=CC(CCC)=C2C(C)=N1
|
|
| InChi Key |
GDFXUTXWCNQTEF-UHFFFAOYSA-N
|
|
| InChi Code |
InChI=1S/C17H19Cl2N7O/c1-4-5-10-6-13(21-16-14(10)9(2)25-26(16)3)23-24-17(27)22-15-11(18)7-20-8-12(15)19/h6-8H,4-5H2,1-3H3,(H,21,23)(H2,20,22,24,27)
|
|
| 化学名 |
1-(3,5-dichloropyridin-4-yl)-3-[(1,3-dimethyl-4-propylpyrazolo[3,4-b]pyridin-6-yl)amino]urea
|
|
| 别名 |
|
|
| HS Tariff Code |
2934.99.9001
|
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
|
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
|
|---|
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.4492 mL | 12.2462 mL | 24.4924 mL | |
| 5 mM | 0.4898 mL | 2.4492 mL | 4.8985 mL | |
| 10 mM | 0.2449 mL | 1.2246 mL | 2.4492 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
|
|---|
 |
 |
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 COA
COA

 463611831
463611831
