| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| Other Sizes |
|

| 靶点 |
KAN0438757 (72 h) reduces the cell viability of Miapaca-2, PANC1, SW620, U-266, and AMO-1 (IC50: 2.75, 3.83, 7.50, 5.08, 11.53 μM, respectively) [1]. KAN0438757 (10 μM, 6 KAN0438757 (50 μM, 12 hours) lowers homologous recombination (HR) activity and increases ionizing radiation (IR)-induced γH2AX foci levels in U2OS cells [1]. KAN0438757 (50 μM, 12 hours ) can reduce HCT-116, SW-1463 and HUVEC KAN0438757 (0-50 μM, 24 hours) can reduce HCT-116 carbohydrates and glycolysis in HUVEC [2].
|
|---|---|
| 体外研究 (In Vitro) |
KAN0438757 (72 h) 降低 Miapaca-2、PANC1、SW620、U-266 和 AMO-1 的细胞活力(IC50 分别为:2.75、3.83、7.50、5.08、11.53 μM)[1]。 KAN0438757(10 μM,6 KAN0438757(50 μM,12 小时)可降低 U2OS 细胞中的同源重组 (HR) 活性并增加电离辐射 (IR) 诱导的 γH2AX 焦点水平 [1]。KAN0438757(50 μM,12 小时)可降低HCT-116、SW-1463 和 HUVEC KAN0438757(0-50 μM,24 小时)可以减少 HUVEC 中的 HCT-116 碳水化合物和糖酵解 [2]。
前药 KAN0438757(为提高细胞通透性和细胞内水解为活性酸形式而设计)在胰腺癌(MIA PaCa-2)、胃癌(NUGC-3)和结肠癌(SW620)细胞系中,能以亚微摩尔级的IC50值显著抑制细胞内果糖-2,6-二磷酸(F-2,6-P2)水平,而其酸形式 KAN0438241 在细胞中的活性较弱 [1]。 通过磺酰罗丹明B法测定,KAN0438757 在长时间(72小时)孵育后能降低多种癌细胞系(MIA PaCa-2, PANC-1, SW620, SW480)的活力 [1]。 在U2OS细胞中,用 KAN0438757 (10 μM)抑制PFKFB3会损害电离辐射(IR)诱导的同源重组(HR)修复因子RPA32和RAD51向核病灶的募集 [1]。 在U2OS细胞的DR-GFP报告基因实验中,KAN0438757 处理将使HR修复活性降低至溶剂对照组的约10%,此效果比ATR抑制更强 [1]。 在IR处理前用 KAN0438757 处理U2OS细胞,会导致IR后24小时γH2AX病灶(DNA双链断裂标志物)水平显著升高,表明DNA修复能力下降 [1]。 KAN0438757 在克隆形成实验中能诱导放射增敏作用。在IR前用 KAN0438757 (10 μM)预处理U2OS细胞,会导致克隆形成呈剂量依赖性减少 [1]。 KAN0438757 的放射增敏作用对转化细胞具有选择性。在同源细胞对中,KAN0438757 联合IR处理能显著降低转化型BJ RAS细胞的克隆形成存活率,而在相同浓度下对非转化型BJ TERT(hTERT永生化)细胞仅产生轻微影响。相比之下,ATM或ATR抑制剂联合IR会严重降低转化和非转化细胞的存活率 [1]。 过表达RAD51可以挽救经 KAN0438757 在其IC50浓度下联合IR处理的BJ RAS细胞的克隆形成存活率,这表明其毒性与其导致的HR功能丧失有关 [1]。 KAN0438757 处理会损害IR诱导的U2OS细胞G2/M期EdU(胸腺嘧啶类似物)掺入增加,提示其在修复合成过程中的脱氧核苷酸掺入中发挥作用 [1]。 KAN0438757 处理消除了IR诱导的PFKFB3与核糖核苷酸还原酶(RNR)RRM2亚基在核病灶中的共定位,并且敲低PFKFB3也会阻断RRM2的募集,这表明PFKFB3的活性位于RRM2向损伤位点募集的上游 [1]。 免疫共沉淀实验显示,在IR后2小时,FLAG标记的PFKFB3与RRM2之间存在相互作用,而在非IR条件下则未检测到 [1]。 KAN0438757 处理(10-30 μM,4小时)会降低U2OS细胞中每个细胞的EdU掺入强度,其效果与羟基脲(HU)相似,表明DNA复制受损 [1]。 DNA纤维实验表明,KAN0438757 处理(10 μM,24小时)严重降低了U2OS细胞中复制叉的速度。这种损害可通过在处理期间补充核苷(腺苷、胞苷、鸟苷、尿苷)来恢复 [1]。 用 KAN0438757 (10 μM)处理U2OS细胞4小时和24小时,会导致所有四种脱氧核苷三磷酸(dNTP)的水平下降50-75% [1]。 与羟基脲(HU)不同,KAN0438757 的抑制不会诱导RPA包裹的ssDNA、p53的诱导或ATR、ATM、Chk1、H2AX的磷酸化,表明它导致复制叉停滞,但并未立即引起复制叉崩溃或检查点激活 [1]。 细胞热转移分析(CETSA)证实了 KAN0438757 在U2OS细胞中与PFKFB3的细胞内靶点结合,其浓度与其细胞活性一致,并且这种结合在处理24小时和72小时后仍然持续 [1]。 |
| 体内研究 (In Vivo) |
在 C57BL6/N 模型中,KAN0438757(腹腔注射,10;25;50 mg/kg)耐受性良好,没有表现出任何显着的全身毒性作用[2]。
|
| 酶活实验 |
使用ADP-Glo法测定重组人PFKFB3的激酶活性。该法通过检测从ATP和果糖-6-磷酸(F6P)生成ADP和F-2,6-P2来量化激酶活性。反应终止后,消耗剩余的ATP。然后将ADP转化回ATP,并通过萤光素酶/萤光素反应测量新合成的ATP,产生的化学发光与激酶活性成正比。抑制剂在与底物孵育前先与酶预孵育 [1]。
使用此实验形式测定了化合物对PFKFB3和PFKFB4的抑制效力(IC50)[1]。 通过将 KAN0438241 (200 μM)滴定到PFKFB3蛋白(20 μM)中进行等温滴定量热法(ITC)实验,以确认结合并确定亲和力 [1]。 |
| 细胞实验 |
克隆形成实验: 将U2OS、BJ TERT或BJ RAS细胞以低密度接种。对于siRNA实验,细胞转染后24小时进行辐照,然后接种进行克隆形成。对于抑制剂研究,细胞用溶剂或 KAN0438757 处理24小时,然后辐照。抑制剂在指定时间后(抑制剂研究为72小时,siRNA研究为4小时)洗去。让细胞形成克隆数天(根据细胞系不同为4-12天),然后固定,用亚甲蓝染色并手动计数 [1]。
免疫荧光显微镜与病灶定量: 将细胞培养在盖玻片上,进行处理和/或辐照。为观察PFKFB3、RPA32、RAD51、γH2AX和RRM2等修复病灶蛋白,细胞在固定前需进行原位细胞分级分离(用含去垢剂的细胞骨架缓冲液提取)以降低背景。然后固定、透化、封闭,并与一抗和荧光二抗孵育。用DAPI染色DNA。通过共聚焦显微镜获取图像。使用CellProfiler软件对病灶数量、强度和共定位进行定量 [1]。 DR-GFP同源重组修复实验: 用siRNA或抑制剂处理U2OS DR-GFP报告细胞。24小时后,转染I-SceI核酸内切酶表达载体以诱导位点特异性双链断裂。转染后24-48小时,收集细胞,通过流式细胞术测量GFP阳性细胞百分比(表示成功的HR修复)。HR活性相对于对照处理细胞计算 [1]。 EdU掺入实验(修复合成): 处理U2OS细胞,辐照,并在不同恢复时间点用EdU脉冲标记30分钟。固定细胞,通过基于点击化学的荧光标记检测掺入的EdU。用Hoechst染色DNA含量。通过流式细胞术定量处于G2/M期(基于DNA含量)且EdU阳性的细胞,以特异性评估修复过程中的核苷酸掺入,排除S期复制 [1]。 EdU掺入实验(复制): 用化合物处理U2OS细胞数小时,在最后40分钟用EdU脉冲标记,然后固定。通过点击化学检测掺入的EdU,并使用图像分析软件(CellProfiler)定量每个细胞的荧光强度 [1]。 DNA纤维实验: 用抑制剂处理U2OS细胞24小时。用两种不同的卤代核苷酸(CldU标记20分钟,然后IdU标记20分钟)顺序标记复制叉。收集细胞,将DNA纤维铺在载玻片上,固定并变性。用特异性抗体检测掺入的核苷酸。使用ImageJ软件测量IdU和CldU轨迹的长度,并计算复制叉速度(单位:千碱基/分钟)[1]。 细胞内dNTP测量: 使用基于HIV-1逆转录酶的方法测量dNTP水平。将细胞胰酶消化、计数,并用甲醇提取dNTP。将干燥的提取物重悬,在存在特定引物/模板和一种dNTP的情况下,由HIV-1 RT进行引物延伸反应。延伸产物的量(对应于提取物中该特定dNTP的水平)被量化。dNTP含量根据细胞数归一化,并与对照处理相比较 [1]。 细胞热转移分析(CETSA): 用 KAN0438757 或DMSO处理U2OS细胞,收集细胞,并通过冻融循环裂解。将细胞裂解物分装,加热到一系列温度,然后离心去除沉淀的蛋白质。通过蛋白质印迹法分析上清液中残留的可溶性PFKFB3蛋白。通过比较药物处理组与溶剂对照组中PFKFB3的溶解曲线来评估其稳定性(指示配体结合)[1]。 Van Schaftingen实验(细胞内F-2,6-P2): 将癌细胞在低葡萄糖/低血清培养基中饥饿过夜,然后用化合物处理并加入葡萄糖刺激。用NaOH裂解细胞,并将裂解液中和。使用基于焦磷酸依赖型磷酸果糖激酶-1(PPi-PFK)被F-2,6-P2强效激活的偶联酶法来量化F-2,6-P2水平。这种激活导致NADH的消耗,通过分光光度法监测。吸光度的下降在线性范围内与F-2,6-P2浓度成正比 [1]。 |
| 药代性质 (ADME/PK) |
在药物发现过程中,化合物的性质被表征,例如缓冲液稳定性、溶解度、代谢稳定性、血浆稳定性、蛋白质结合和渗透性[1]。
具体而言,KAN0438757(活性酸KAN0438241的酯)的前药设计旨在提高细胞渗透性,并实现有效的细胞内水解以释放活性化合物[1]。 |
| 毒性/毒理 (Toxicokinetics/TK) |
与癌细胞系不同,外周血单核细胞 (PBMC) 的活力在 PFKFB3 被 KAN0438757 抑制后并未受到影响 [1]。
在克隆形成实验中,未转化的 BJ TERT 细胞在 KAN0438757 浓度下仅表现出轻微的存活率下降,而转化的 BJ RAS 细胞的存活率却显著降低,尤其是在与 IR 联合使用时,这表明存在潜在的治疗窗口 [1]。 |
| 参考文献 | |
| 其他信息 |
KAN0438757是一种选择性小分子糖酵解酶PFKFB3抑制剂,其开发目的是为了研究PFKFB3在DNA修复中的作用[1]。
该研究揭示了PFKFB3在DNA损伤反应中一种新的、非经典的作用。PFKFB3通过MRN-ATM-γH2AX-MDC1通路被募集到DNA双链断裂位点。其酶活性对于后续募集核糖核苷酸还原酶的 RRM2 亚基和 HR 修复因子(RPA、RAD51)是必需的,从而促进局部 dNTP 的产生和修复合成 [1]。 用 KAN0438757 抑制 PFKFB3 会损害同源重组修复,减少细胞 dNTP 池,停滞复制叉,并使癌细胞对电离辐射敏感,同时不影响非转化细胞。这使得PFKFB3抑制成为癌症特异性放射增敏的潜在策略[1]。 先前报道的PFKFB3抑制剂3-PO在作者的酶促或细胞HR修复试验中未显示活性,促使人们开发了KAN0438757[1]。 活性代谢物KAN0438241与人PFKFB3的共晶结构显示,它结合在果糖-6-磷酸底物口袋中,模拟内源性底物的相互作用,并且是一种非ATP竞争性抑制剂[1]。 |
| 分子式 |
C21H18FNO7S
|
|---|---|
| 分子量 |
447.433528423309
|
| 精确质量 |
447.078
|
| CAS号 |
1451255-59-6
|
| PubChem CID |
71586631
|
| 外观&性状 |
Typically exists as solid at room temperature
|
| LogP |
3.2
|
| tPSA |
142
|
| 氢键供体(HBD)数目 |
4
|
| 氢键受体(HBA)数目 |
9
|
| 可旋转键数目(RBC) |
8
|
| 重原子数目 |
31
|
| 分子复杂度/Complexity |
700
|
| 定义原子立体中心数目 |
0
|
| SMILES |
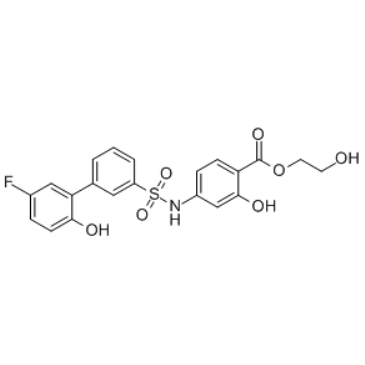
C1=CC(=CC(=C1)S(=O)(=O)NC2=CC(=C(C=C2)C(=O)OCCO)O)C3=C(C=CC(=C3)F)O
|
| InChi Key |
QRDFCYMUXHUTCS-UHFFFAOYSA-N
|
| InChi Code |
InChI=1S/C21H18FNO7S/c22-14-4-7-19(25)18(11-14)13-2-1-3-16(10-13)31(28,29)23-15-5-6-17(20(26)12-15)21(27)30-9-8-24/h1-7,10-12,23-26H,8-9H2
|
| 化学名 |
2-hydroxyethyl 4-[[3-(5-fluoro-2-hydroxyphenyl)phenyl]sulfonylamino]-2-hydroxybenzoate
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO : ~130 mg/mL (~290.55 mM)
|
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: 2.17 mg/mL (4.85 mM) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液; 超声助溶。
例如,若需制备1 mL的工作液,可将100 μL 21.7 mg/mL澄清DMSO储备液加入400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: ≥ 2.17 mg/mL (4.85 mM) (饱和度未知) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 例如,若需制备1 mL的工作液,可将 100 μL 21.7 mg/mL澄清DMSO储备液加入900 μL 20% SBE-β-CD生理盐水溶液中,混匀。 *20% SBE-β-CD 生理盐水溶液的制备(4°C,1 周):将 2 g SBE-β-CD 溶解于 10 mL 生理盐水中,得到澄清溶液。 View More
配方 3 中的溶解度: ≥ 2.17 mg/mL (4.85 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.2350 mL | 11.1749 mL | 22.3499 mL | |
| 5 mM | 0.4470 mL | 2.2350 mL | 4.4700 mL | |
| 10 mM | 0.2235 mL | 1.1175 mL | 2.2350 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
 Tepotinib hydrochloride monohydrate
Tepotinib hydrochloride monohydrate
 BKN-1
BKN-1
 NEO214
NEO214
 MJO445
MJO445
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 463611831
463611831
