| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 1mg |
|
||
| 5mg |
|
||
| 10mg |
|
||
| Other Sizes |
|

| 靶点 |
Acetyl-CoA targets a wide range of enzymes and transporters that utilize it as a substrate or ligand. Its targets include: (1) Acetyl-CoA Carboxylase (ACC): Acetyl-CoA serves as the substrate for this enzyme to synthesize malonyl-CoA ; (2) Citrate Synthase: In the TCA cycle, acetyl-CoA condenses with oxaloacetate catalyzed by this enzyme to form citrate ; (3) SLC33A1 (AT-1): This is the only identified acetyl-CoA transmembrane transporter, located on the ER membrane, responsible for translocating acetyl-CoA from the cytosol into the ER lumen ; (4) NLRX1: A mitophagy receptor. Cytosolic acetyl-CoA acts as a signaling metabolite that directly binds to NLRX1 to regulate nutrient starvation-induced mitophagy. In vitro binding assays showed that unlabeled acetyl-CoA competes with biotin-acetyl-CoA for binding to recombinant NLRX1 protein with a saturation concentration of approximately 25 μM .
; Endogenous Metabolite
|
|---|---|
| 体外研究 (In Vitro) |
在饥饿的 U2OS 细胞中,乙酰辅酶 A 三钠促进细胞质蛋白乙酰化,同时减少饥饿诱导的自噬通量。 (稳定表达 GFP-LC3 的 U2OS 细胞显微注射乙酰辅酶 A 三钠;然后在缺乏营养物的情况下用 100 nM BafA1 培养,三小时后固定)[2]。
- 在饥饿的U2OS细胞中,显微注射乙酰辅酶A可减少饥饿诱导的自噬通量,同时增加细胞质蛋白的乙酰化水平。 - 在稳定表达GFP-LC3的U2OS细胞中,使用轻度饥饿培养基(含5 mM葡萄糖和2 mM谷氨酰胺)处理可降低胞质乙酰辅酶A水平,并显著降低线粒体蛋白TIM23、MT-CO2和HSP60的水平,同时增加LC3向线粒体的招募,表明线粒体质量减少源于自噬降解。 - 在KRAS突变的肿瘤细胞(如KPC细胞和AsPC-1细胞)中,使用KRAS抑制剂(MRTX1133和RMC-6236)处理可显著降低ACLY表达及胞质乙酰辅酶A水平,并诱导线粒体自噬,该效应可被补充乙酸所阻断。 |
| 体内研究 (In Vivo) |
- 在体内,乙酰辅酶A可防止小鼠因压力超负荷而患上心肌病。
- 在小鼠实验中,禁食一天会导致心脏和肌肉等器官中总乙酰辅酶A水平显著降低,这与较低的蛋白质乙酰化水平相关;然而,在相同实验条件下,肝脏的蛋白质乙酰化和乙酰辅酶A水平却升高,而对大脑乙酰辅酶A浓度无明显影响。 - 在KRAS抑制剂耐药机制研究中,使用体内移植瘤模型发现,NLRX1敲除可显著增强KRAS抑制剂MRTX1133对KPC肿瘤的抑制效果(表现为肿瘤体积和重量减小),同时肿瘤组织中活性氧水平升高而线粒体自噬受损,表明KRAS抑制剂通过诱导NLRX1依赖的线粒体自噬帮助肿瘤细胞抵抗氧化应激而产生耐药。 在小鼠心脏压力超负荷模型中,乙酰辅酶 A 三钠通过抑制适应不良的自噬来减轻压力超负荷引起的心肌病[2][3]。小鼠一整天不进食(但允许无限量饮水),其心脏和肌肉中总乙酰辅酶 A 三钠的水平显着降低,这与较低水平的蛋白质乙酰化相关。然而,相同的实验设置实际上提高了蛋白质乙酰化和乙酰辅酶 A 三钠的肝脏水平,同时对大脑中酶的浓度没有明显影响[4]。 |
| 酶活实验 |
- 乙酰辅酶A羧化酶(ACC)活性检测(NADH速率法):该原理基于ACC催化乙酰辅酶A、NaHCO3和ATP生成丙二酰辅酶A、ADP和无机磷。通过偶联丙酮酸激酶和乳酸脱氢酶系统,将生成的ADP和磷酸烯醇式丙酮酸转化为丙酮酸,进一步将NADH氧化为NAD+。由于NADH在340 nm处具有特征吸收峰,通过监测340 nm处吸光值的下降速率即可计算ACC的活性。
- 乙酰辅酶A羧化酶活性检测(非放射性分光光度法):将透化处理后的谷氨酸棒杆菌细胞加入含有乙酰辅酶A、碳酸氢盐、镁离子和ATP的反应混合物中。在设定的时间点取出等分试样,加入三氟乙酸终止反应。通过柠檬酸合酶法测定每个等分试样中剩余的乙酰辅酶A水平,该方法通过分光光度法在412 nm处监测二硫代双硝基苯甲酸-硫酚盐黄色化合物的形成。 - NLRX1与乙酰辅酶A直接结合实验(下拉实验):使用昆虫细胞表达系统纯化获得N端MBP标签的重组NLRX1蛋白。体外下拉实验中,将重组MBP-NLRX1蛋白与生物素标记的乙酰辅酶A孵育,通过链霉亲和素磁珠捕获复合物,洗脱后通过Western blot检测MBP-NLRX1。未标记的乙酰辅酶A可以剂量依赖方式竞争生物素-乙酰辅酶A与重组NLRX1蛋白的结合,竞争饱和浓度约为25 μM。 |
| 细胞实验 |
- 细胞培养与饥饿处理:将U2OS细胞(稳定表达GFP-LC3)在常规培养基中培养。为诱导自噬,将细胞换入轻度饥饿培养基(含5 mM葡萄糖和2 mM谷氨酰胺),处理指定时间(如3小时)。为阻断自噬流,可在处理前加入巴弗洛霉素A1 (BafA1, 100 nM)。处理结束后,细胞用PBS洗涤,裂解后通过Western blot检测线粒体蛋白(如TIM23、MT-CO2、HSP60)水平,或通过荧光显微镜观察GFP-LC3斑点形成。
- 乙酰辅酶A显微注射:在饥饿的U2OS细胞中,通过显微注射将乙酰辅酶A直接导入细胞质。注射后,细胞在无营养条件下用BafA1 (100 nM) 处理3小时,然后固定。通过检测GFP-LC3斑点评估自噬通量,同时检测细胞质蛋白乙酰化水平。 - KRAS抑制剂处理:将KRAS突变的肿瘤细胞(如KPC细胞或AsPC-1细胞)接种于培养板。使用KRAS抑制剂MRTX1133或RMC-6236处理细胞。同时设置对照组(DMSO)和乙酸补充组(以验证乙酰辅酶A的特异性作用)。处理结束后,收集细胞裂解液,通过Western blot检测ACLY表达水平、线粒体蛋白水平及乙酰辅酶A水平。 |
| 动物实验 |
- 心肌病模型:通过压力超负荷诱导小鼠心肌病模型,给予乙酰辅酶A治疗,评估其对心肌病的预防效果。
- 禁食实验:将小鼠分为自由进食组和禁食组(禁食24小时,不禁水)。禁食结束后,处死小鼠,收集心脏、肌肉、肝脏和脑组织。通过LC-MS等方法检测各组织中总乙酰辅酶A水平,并通过Western blot检测蛋白质乙酰化水平。 - 肿瘤移植瘤模型:将KPC细胞(KRAS突变胰腺癌细胞)皮下接种于免疫缺陷小鼠体内,建立移植瘤模型。待肿瘤生长至一定体积后,将小鼠随机分为对照组和给药组(如KRAS抑制剂MRTX1133),部分实验使用NLRX1敲除的KPC细胞。通过口服灌胃或腹腔注射给予KRAS抑制剂。给药期间定期测量肿瘤体积和小鼠体重。实验结束后处死小鼠,剥离肿瘤组织称重,检测肿瘤组织中活性氧水平和线粒体自噬相关指标。 |
| 药代性质 (ADME/PK) |
乙酰辅酶A是一种高极性、带电荷的分子(分子量约800),无法自由扩散通过细胞膜,需要特异性转运蛋白(如SLC33A1)介导其跨膜转运。在实验室条件下,乙酰辅酶A水溶液可在-20°C保存长达2周。
|
| 毒性/毒理 (Toxicokinetics/TK) |
文献未描述乙酰辅酶A本身的直接毒性信息。作为内源性代谢物,其在生理浓度下不表现毒性。根据供应商信息,乙酰辅酶A锂盐三水合物在运输中未被归类为危险物质。需要注意的是,乙酰辅酶A稳态的失衡与多种疾病相关,包括神经退行性疾病、衰老相关疾病和癌症等。例如,SLC33A1的功能缺失或过度活跃均可导致疾病,尤其是影响神经系统的疾病。
|
| 参考文献 | |
| 其他信息 |
乙酰辅酶A参与脂肪酸和甾醇的生物合成、脂肪酸的氧化以及多种氨基酸的代谢。它还作为一种生物乙酰化试剂发挥作用。
赖氨酸乙酰化是一种保守的蛋白质翻译后修饰,它将乙酰辅酶A的代谢与细胞信号传导联系起来。近年来,质谱技术在赖氨酸乙酰化的鉴定和定量方面取得了显著进展,加深了我们对赖氨酸乙酰化的理解,揭示了其通过调控蛋白质相互作用、活性和定位参与多种生物学过程。此外,蛋白质还经常发生其他类型的酰化修饰,例如甲酰化、丁酰化、丙酰化、琥珀酰化、丙二酰化、肉豆蔻酰化、戊二酰化和巴豆酰化。赖氨酸酰化与细胞代谢之间错综复杂的联系已通过多种代谢物敏感的酰化反应及其被sirtuin脱酰酶选择性去除而得以阐明。这些新发现揭示了不同赖氨酸酰化和脱酰酶的新功能,同时也突显了乙酰化调控各种细胞过程的机制。[1]乙酰辅酶A (AcCoA) 是脂肪、糖和蛋白质分解代谢交汇处营养状态的重要整合因子。本文研究表明,营养匮乏会导致AcCoA快速消耗。AcCoA的消耗导致胞质蛋白整体乙酰化水平相应降低,并诱导自噬——一种维持细胞稳态的自我消化过程。多种旨在增加或减少胞质乙酰辅酶A(AcCoA)水平的不同操作分别导致培养的人类细胞和小鼠体内自噬的抑制或诱导。此外,在心脏压力负荷过重模型中,维持高水平的AcCoA可抑制适应不良的自噬。AcCoA的耗竭降低了乙酰转移酶EP300的活性,而EP300是高水平AcCoA抑制自噬所必需的。总之,我们的结果表明胞质AcCoA作为自噬的核心代谢调节因子发挥作用,从而阐明了以AcCoA为中心的药理学策略,可用于治疗性地调控自噬。[2]心脏肥大是心力衰竭的主要预测因子,也是一种高死亡率的常见疾病。然而,对于从稳定型心脏肥大到失代偿性心力衰竭的转变机制知之甚少。本研究探讨了自噬的作用。自噬是一种保守的通路,介导长寿命蛋白和细胞器的大量降解,最终可能导致细胞死亡。为了量化自噬活性,我们构建了“自噬报告基因”小鼠品系,并证实短期营养缺乏可在体内诱导心肌细胞自噬。主动脉缩窄引起的压力负荷可导致心力衰竭,并显著增强心脏自噬。负荷诱导的自噬活性在48小时达到峰值,并至少持续3周显著升高。此外,自噬活性并非空间均匀分布,而是在基底间隔中尤为显著。编码Beclin 1(一种早期自噬体形成所必需的蛋白)的基因杂合性缺失可降低心肌细胞自噬,并减弱严重压力应激诱导的病理性重塑。相反,Beclin 1过表达可增强自噬活性,并加剧病理性重塑。综上所述,这些研究结果表明自噬参与了负荷诱导性心力衰竭的发病机制,并提示其可能成为新型治疗干预的靶点。[3]乙酰辅酶A(乙酰CoA)是一种重要的代谢中间体。不同亚细胞区室中乙酰CoA的丰度反映了细胞的整体能量状态。此外,乙酰CoA的浓度通过变构调节或改变底物可用性来影响多种酶的活性或特异性。最后,乙酰CoA通过影响包括组蛋白在内的多种蛋白质的乙酰化谱,直接或通过表观遗传调控基因表达来控制关键的细胞过程,包括能量代谢、有丝分裂和自噬。因此,乙酰CoA通过同时作为代谢中间体和第二信使发挥作用,决定了细胞分解代谢和合成代谢之间的平衡。[4] |
| 分子式 |
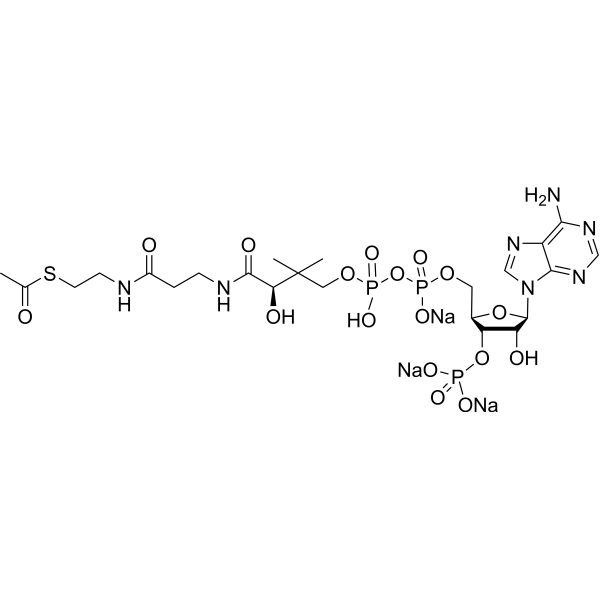
C23H35N7NA3O17P3S
|
|---|---|
| 分子量 |
875.52
|
| 精确质量 |
809.125
|
| CAS号 |
102029-73-2
|
| 相关CAS号 |
Acetyl coenzyme A lithium;32140-51-5;Acetyl coenzyme A;72-89-9
|
| PubChem CID |
6302
|
| 外观&性状 |
White to off-white solid powder
|
| 密度 |
1.9±0.1 g/cm3
|
| 折射率 |
1.718
|
| LogP |
-3.89
|
| tPSA |
418.36
|
| 氢键供体(HBD)数目 |
9
|
| 氢键受体(HBA)数目 |
22
|
| 可旋转键数目(RBC) |
20
|
| 重原子数目 |
51
|
| 分子复杂度/Complexity |
1380
|
| 定义原子立体中心数目 |
4
|
| SMILES |
O[C@@H]1[C@@H]([C@@H](COP(O)(=O)OP(O)(=O)OCC(C)(C)[C@@H](O)C(=O)NCCC(=O)NCCSC(=O)C)O[C@H]1N1C=NC2C(=NC=NC1=2)N)OP(O)(O)=O.[Na+].[Na+].[Na+]
|
| InChi Key |
ZSLZBFCDCINBPY-KMYLAXNMSA-N
|
| InChi Code |
InChI=1S/C23H38N7O17P3S/c1-12(31)51-7-6-25-14(32)4-5-26-21(35)18(34)23(2,3)9-44-50(41,42)47-49(39,40)43-8-13-17(46-48(36,37)38)16(33)22(45-13)30-11-29-15-19(24)27-10-28-20(15)30/h10-11,13,16-18,22,33-34H,4-9H2,1-3H3,(H,25,32)(H,26,35)(H,39,40)(H,41,42)(H2,24,27,28)(H2,36,37,38)/t13-,16-,17-,18?,22-/m1/s1
|
| 化学名 |
S-[2-[3-[[4-[[[(2R,3S,4R,5R)-5-(6-aminopurin-9-yl)-4-hydroxy-3-phosphonooxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-hydroxyphosphoryl]oxy-2-hydroxy-3,3-dimethylbutanoyl]amino]propanoylamino]ethyl] ethanethioate
|
| 别名 |
Acetyl coenzyme A sodium salt; 102029-73-2; Coenzyme A, S-acetate, trisodium salt (9CI); Acetyl Coenzyme A Trisodium Salt; Acetyl-CoA sodium; XGCDESRMLMCTPI-UHFFFAOYSA-N;
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month 注意: 请将本产品存放在密封且受保护的环境中,避免吸湿/受潮。 |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
H2O : 83.33 mg/mL (95.18 mM)
DMSO : < 1 mg/mL |
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: 100 mg/mL (114.22 mM) in PBS (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液; 超声助溶。
请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 1.1422 mL | 5.7109 mL | 11.4218 mL | |
| 5 mM | 0.2284 mL | 1.1422 mL | 2.2844 mL | |
| 10 mM | 0.1142 mL | 0.5711 mL | 1.1422 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
 L-Histidinol-d3
L-Histidinol-d3
 D-myo-Inositol-3-phosphate sodium
D-myo-Inositol-3-phosphate sodium
 (±)19(20)-DiHDPA
(±)19(20)-DiHDPA
 4-Hydroxy-2-butanone
4-Hydroxy-2-butanone
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 463611831
463611831
