| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 1mg |
|
||
| 2mg |
|
||
| 5mg |
|
||
| 10mg |
|
||
| Other Sizes |
|
| 靶点 |
DNA/RNA synthesis; Mycotoxin
|
|---|---|
| 体外研究 (In Vitro) |
Deoxynivalenol/脱氧雪腐镰刀菌烯醇(DON)是一种霉菌毒素的毛霉烯家族,其人类暴露水平可能很高。流行病学研究表明DON与胃肠道疾病之间存在联系。我们研究了DON与Caco-2细胞的相互作用,Caco-2细胞是一种广泛用于人体肠道屏障的体外模型。DON的根尖到基底外侧(吸收)和基底外侧到根尖(排泄)的运输与初始浓度和孵育时间严格成正比。吸收和排泄的平均速率与甘露醇相似,并且在EGTA(一种钙螯合剂)的存在下增加。这些数据表明DON通过紧密连接通过细胞旁途径穿过肠粘膜,尽管不排除一些被动的跨细胞扩散。DON转运不受p -糖蛋白(PgP)或多药耐药相关蛋白(MRPs)抑制剂的影响。长期暴露于DON可引起丝裂原活化蛋白激酶(MAPKs) Erk1/2、p38和SAPK/JNK的磷酸化,并降低经上皮抵抗,提示DON可引发肠道炎症。这些数据表明,长期暴露于DON污染的食物可能会通过改变肠粘膜完整性和诱导与炎症有关的MAPKs对人体健康产生负面影响。[1]
|
| 体内研究 (In Vivo) |
Deoxynivalenol/脱氧雪腐镰刀菌醇(DON, vomitoxin)是世界范围内谷物中最常见的污染物之一。研究了DON对Charles River Sprague-Dawley大鼠胎儿发育的影响。妊娠第6-19天(GD),以0、0.5、1、2.5或5 mg/kg体重剂量的DON灌胃妊娠雌性大鼠,每天1次。妊娠第20天剖宫产时,测定生殖发育指标。所有雌性存活至剖宫产。DON引起了怀孕雌性小鼠过量流涎的剂量相关增加,这种反应可能与大鼠缺乏呕吐反射有关。在5 mg/kg剂量下,全孕期采食量和平均增重显著降低,平均增重(胴体重)和妊娠子宫重显著降低,52%的窝仔(12/23)被完全吸收,平均每窝早、晚死亡数显著增加,平均胎重和冠臀长显著降低,小不点发生率显著增加,胎儿胸骨骨化率显著降低。中央、背弓、椎骨、跖骨和掌骨明显减少。在2.5 mg/kg剂量下,DON显著降低胎儿平均体重、冠臀长和椎体骨化。这些影响可能继发于母体毒性和胎儿体积缩小。在5.0 mg/kg剂量下,胸骨不对准和融合的发生率显著增加。0.5和1.0 mg/kg剂量对发育无不良影响。在所有治疗组中均观察到与剂量相关的母体肝脏体重/体重比增加(在1、2.5和5 mg/kg时显著增加)。体重变化与剂量相关的肝细胞细胞质改变相关。本研究中母体毒性的NOEL为0.5 mg/kg,基于肝-体重比在1 mg/kg时的剂量相关增加。胎儿毒性的下限是1毫克/公斤,这是基于2.5毫克/公斤和5毫克/公斤的胎儿发育一般降低。在Sprague-Dawley大鼠胸骨发育异常的基础上,DON被认为是一种致畸剂,剂量为每天5毫克/公斤。[3]
脱氧雪腐镰刀菌烯醇(DON或呕吐毒素)是一种常见于谷物中的毛霉毒素,对实验动物的生长和免疫功能产生不利影响。采用竞争性酶联免疫吸附试验(ELISA)监测口服25mg/kg bw DON毒素的年轻成年B6C3F1雄性小鼠组织中DON的分布和清除动力学。血浆、肝脏、脾脏和脑组织在5 ~ 24h检测到DON,心脏和肾脏在5 ~ 8h检测到DON。给药后5-15分钟(12微克/毫升)内观察到最高的DON血浆浓度。两室动力学(t(1/2) α =20.4 min, t(1/2) β =11.8h)快速清除,8h和24h后血浆DON最大浓度分别为5%和2%。DON在其他组织中的分布和清除动力学与血浆相似。5 min时,DON浓度(杯/g)肝脏为19.5+/-1.9,肾脏为7.6+/-0.5,脾脏为7.3+/-0.8,心脏为6.8+/-0.9,大脑为0.8+/-0.1。酶联免疫吸附法在组织中的DON回收率与先前使用(3)H-DON和25mg/kg bw DON剂量的研究相当。该酶联免疫吸附试验进一步适用于通过饮食接触毒素的小鼠血浆中DON的检测。该方法提供了一种简单的策略,可用于回答啮齿动物中剂量、物种、年龄、性别、遗传背景和暴露途径/持续时间如何影响DON摄取和清除的相关问题。[5] 脱氧雪腐镰刀菌烯醇 (DON) 是一种单端孢霉烯族霉菌毒素,对实验动物的生长和免疫功能有不利影响。低剂量可快速、短暂地上调促炎细胞因子(如 IL-1β、IL-6、TNF-α),导致免疫刺激,而高剂量则引起淋巴组织细胞凋亡,导致免疫抑制。[5] 在小鼠中,急性口服暴露于 DON (25 mg/kg 体重) 会在脾脏中依次诱导:15-30 分钟内 JNK 1/2、ERK 1/2 和 p38 的磷酸化;1-2 小时内转录因子的激活;以及 1-4 小时内细胞因子 mRNA 的表达。[5] DON 还具有神经内分泌效应,包括厌食和呕吐,这可能由血清素能系统介导。[5] |
| 酶活实验 |
转运研究[1]
在运输实验中,将培养基替换为含有5 mM葡萄糖和10 mM Hepes (pH 7.4)的汉克平衡盐溶液(HBSS),用于插入物的下腔室,或10 mM Mes (pH 6.0)用于上腔室。下腔室进一步添加1% (w/v)牛血清白蛋白。脱氧雪腐镰梨醇(DON)或4.25 μM [14C]甘露醇分别加入1.8 ml或2.8 ml的输送介质。在动力学实验中,以2 μg/ml的浓度在供体室中引入Deoxynivalenol (DON)),在剂量效应实验中,以0.16 ~ 7.5 μg/ml的不同浓度引入Deoxynivalenol (DON)。在每次取样时,从接受室中取出等量的输送介质,并用新鲜的输送介质代替。样品在2ml Aqualuma®中分散后,用高效液相色谱法分析样品Deoxynivalenol (DON)含量,用液体闪烁光谱法分析样品的[14C]甘露醇含量。为了检测可能出现的Deoxynivalenol (DON)-共轭代谢物,一些样品在pH为5的0.2 M醋酸缓冲液中,在37°C下用β-葡萄糖醛酸酶和芳基硫酸酯酶的混合物处理24 h,然后进行HPLC分析。 MAPKs磷酸化的评估[1] 细胞在微孔膜上培养21-27天,如运输研究所述。然后在不含EGF和胰岛素的相同培养基中培养过夜。然后以2 μg/ml的浓度在上隔室中添加Deoxynivalenol (DON),持续指定时间或在24 h内以不同浓度添加。 碱性磷酸酶(AP)活性[2] 在培养22 d内,通过形成蓝色双双胍沉淀来检测碱性磷酸酶的细胞酶活性。 以240 μL原液A (25 mg/mL硝基蓝氯化四氮唑[NBT]在70%二甲基甲酰胺[DMF]中)和60 μL B (50 mg/mL 5-溴-4-氯-3-吲哚磷酸甲苯胺盐[BCIP]在100% DMF中)在16 mL 0.1 M Tris-HCl中配制成工作溶液。用200 μL的工作液孵育孔,在黑暗中过夜,然后去除工作液,在200 μL DMF中溶解双双胍3 h。在560nm处测定吸光度,并采用校准曲线计算双双胍的吸光度。 竞争直接ELISA法测定脱氧雪腐镰刀菌醇(DON) [5] 脱氧雪腐血毒素(DON)使用商业Deoxynivalenol (DON) ELISA进行分析,该ELISA基于先前描述的直接竞争ELISA (Casale等,1988)。本实验中使用的单克隆抗体与DON和3-ADON结合,但不与其他毛链烯结合。由于该试验旨在分析谷物和谷物产品中的DON,因此制造商的方案适用于血浆和器官组织样品,通过在10% (v/v)的人血浆或1%牛血清白蛋白(w/v)的PBS中制备单独的DON标准品(4至1,000 ng/ml),以防止非特异性结合。样品或标准品(各100 μl)与100 μl DON辣根过氧化物酶偶联液在生产厂家提供的混合孔中混合。取该混合物的100 μl转移到抗体包被的孔中,在25℃下孵育5 min。洗孔,加入100 μl K-blue Max TMB底物,在25℃下孵育5 min。用100 μl 2N硫酸停止反应,在Molecular Devices ELISA阅读器上用450 nm滤光片读取吸光度。使用Molecular Devices Softmax软件从标准曲线定量DON浓度。在初步研究中观察到90%或更高的DON回收率,在加热的肝脏提取物中直接加入12.5至250 ng/g组织中的毒素并进行分析,或者在肝脏匀浆中加入10至250 ng/g组织中的毒素,加热并分析上清。由于该ELISA可以检测DON及其一些代谢物,因此报告的数据为每ml血浆或每g器官组织的DON当量。 |
| 细胞实验 |
细胞毒性测定[1]
采用比色法3-(4,5二甲基噻唑-2-基)-2,5-二苯基溴化四氮唑(MTT)测定Deoxynivalenol (DON)对Caco-2细胞增殖的影响。细胞在96孔板(Nunc, Roskilde, Denmark)上接种,密度为35000个细胞/cm2,接种24 h后,用Deoxynivalenol (DON),浓度在0 ~ 10 μg/ml之间,孵育48 h。MTT实验与Mosmann(1983)一样,使用100 μl MTT (0.5 mg/ml PBS), 37℃孵育2 h,晶体在100 μl DMSO中溶解,在500 nm处读取。 在MTT实验结束或运输实验结束时,用LDH法测定脱氧雪腐镰刀菌醇(DON)的细胞毒作用,LDH法作为试剂盒购买。将细胞暴露于1% (v/v) Triton X-100中获得最大LDH释放。在500 nm处测定了还原甲醛的反应产物。 凋亡测定[1] 细胞死亡检测ELISA [1] 采用细胞死亡检测elisa +试剂盒检测DNA片段,检测凋亡细胞的死亡情况。细胞以35000个细胞/cm2的密度接种于96孔板,接种24 h后,用Deoxynivalenol (DON)在30、90和300 ng/ml浓度下孵育48 h。用丁酸钠在50 mM浓度条件下孵育Deoxynivalenol (DON)作为阳性对照。细胞测定按照制造商的说明进行。[1] 凋亡细胞的荧光显微镜检测[1] 荧光显微镜下吖啶橙/碘化丙啶染色观察细胞核形态。以35000个细胞/cm2的密度在BD Falcon™CultureSlides中接种Caco-2细胞,在播种后24小时,在30、90和300 ng/ml浓度下,用Deoxynivalenol (DON)孵育48小时。在相同条件下,用丁酸钠孵育50 mM的细胞作为阳性对照。然后用吖啶橙(1mg /ml)/碘化丙啶(88 μg/ml)的混合物对细胞进行短暂处理。荧光显微镜下计数凋亡细胞(细胞核高度凝聚或碎片化)和非凋亡细胞(细胞核完整)。 测定96孔板格式[2] 采用96孔板法分析细胞活力(细胞计数、乳酸脱氢酶(LDH)测定、中性红(NR)摄取、MTT测定)、增殖(BrdU测定)和凋亡(发光caspase 3/7测定)。将IPEC-1和IPEC-J2细胞接种于96孔塑料组织培养板,培养4 d至汇合。取出培养基,用PBS洗涤一次后,加入含有Deoxynivalenol (DON) (100-4000 ng/mL)终浓度增加的新鲜培养基。细胞孵育24小时、48小时或72小时。对于长期实验(14天),以相同的方式进行处理,但使用较低的Deoxynivalenol (DON)浓度(50-500 ng/mL),并在每3-4天后定期交换培养基+ Deoxynivalenol (DON)。所有实验均进行三次重复,并使用多板读取器进行至少三次独立实验。 流式细胞术分析细胞周期[2] 将IPEC-1和IPEC-J2细胞接种于塑料组织培养6孔板中,用融合细胞层(4 d)进行实验。然后将细胞同步于无FCS培养基中24 h,并按上述方法暴露于Deoxynivalenol (DON)。细胞胰蛋白酶化、成球并在PBS中重悬。乙醇固定和碘化丙啶(PI)染色程序如前所述进行。 |
| 动物实验 |
雌性适应环境后进行交配(每只雄性配两只雌性)。交配日当天下午约4:30开始同居。次日早晨,检查每只雌性阴道冲洗液中是否存在精子。任何精子阳性的雌性均被假定为怀孕(妊娠第0天),并采用分层随机化方法将每只动物分配到对照组或四个处理组之一。五个处理组,每组24只雌性,在妊娠第6-19天通过灌胃给予0、0.5、1.0、2.5或5.0 mg/kg体重/天的脱氧雪腐镰刀菌烯醇(DON)。在处理期间和妊娠第20天,每天测量饲料和饮水消耗量以及体重。每天称量雌性体重,并根据每只动物的体重确定给予的水或脱氧雪腐镰刀菌烯醇(DON)溶液的体积。脱氧雪腐镰刀菌烯醇 (DON) 的最大给药体积为 1 ml/100 g 体重。[3]
本研究的剂量基于一项重复剂量范围探索研究的毒性数据,该研究在妊娠第 6-19 天 (GD 6-19) 对怀孕雌性大鼠灌胃给予 0.25 至 7.5 mg/kg 体重的脱氧雪腐镰刀菌烯醇 (DON)。7.5 mg/kg 体重的剂量具有胎儿毒性,因此未在本研究中使用。在剂量范围探索研究中,适应性饲养、饲养条件、交配、动物处理、饲料、饮水、化合物给药、胎儿分析等均与本文报道的研究相同。[3] 在急性暴露研究中,在给予脱氧雪腐镰刀菌烯醇 (DON) 前 1 小时停止喂食和饮水。将脱氧雪腐镰刀菌烯醇 (DON) 以 25 mg/kg 体重的剂量,溶于 250 μl 无内毒素水中,通过灌胃法给予小鼠。灌胃后立即恢复小鼠的食物和饮水。未灌胃的小鼠作为对照组。分别在 5、15、30 分钟以及 1、2、4、8、24 小时后,对小鼠进行麻醉并采集血液。随后处死小鼠,取出肝脏、肾脏、心脏、脾脏和脑组织。分离血浆并储存于 -20°C。将各器官冷冻于 -80°C,用研钵和研杵研磨成粉末,然后与磷酸盐缓冲液 (PBS) 按 1:5 的比例混合。将组织提取物以 14,000 × g 的转速离心 10 分钟。将所得上清液加热至 100°C 5 分钟,以灭活酶并沉淀蛋白质。将加热后的提取物以 14,000 × g 离心 10 分钟,然后将上清液适当稀释后用于 ELISA 检测。[5] 在亚慢性喂养研究中,按照先前描述的方法(Pestka 等,1989),将纯化的脱氧雪腐镰刀菌烯醇 (DON) 以 0、2、5、10 和 20 mg/kg 的浓度添加到粉状高脂 AIN-93 G 纯化啮齿动物饲料 101847 中。实验饲料置于专用容器中,以最大程度地减少洒落。实验期间,笼子放置在 II 级通风柜中。小鼠饲喂4周后,按照先前描述的方法(Hem等,1998)从隐静脉采集血液。[5] 药代动力学分析[5] 采用双室开放模型(Shargel等,2004)计算毒代动力学参数。血浆和组织中脱氧雪腐镰刀菌烯醇(DON)的浓度采用双指数表达式拟合,以计算清除率。[5] 急性口服暴露研究:雄性B6C3F1小鼠(7周龄)在给药前禁食1小时。将脱氧雪腐镰刀菌烯醇溶解于无内毒素水中,以25 mg/kg体重的剂量,通过灌胃法给药,灌胃体积为250 μL。灌胃后立即恢复食物和水的供应。给药后5、15、30分钟以及1、2、4、8、24小时,对小鼠进行麻醉,采集血液,然后处死小鼠。取出肝脏、肾脏、心脏、脾脏和脑组织。分离血浆并储存于-20℃。将各器官冷冻于-80℃,研磨成粉末,以1:5的比例与磷酸盐缓冲液(PBS)匀浆,离心,取上清液于100℃加热5分钟以灭活酶并沉淀蛋白质。再次离心后,取上清液进行ELISA分析。 [5]亚慢性喂养研究:将纯化的脱氧雪腐镰刀菌烯醇(DON)以0、2、5、10和20 mg/kg的浓度混入高脂AIN-93G纯化啮齿动物饲料粉中。小鼠饲喂这些饲料4周。从隐静脉采集血液进行血浆DON分析。[5] |
| 药代性质 (ADME/PK) |
吸收、分布和排泄
已在动物身上开展了代谢研究,主要使用T-2毒素,少数使用脱氧雪腐镰刀菌烯醇(DON)。这些单端孢霉烯族毒素能迅速从消化道吸收……毒素分布相当均匀,不会在任何特定器官或组织中明显蓄积。单端孢霉烯族毒素可通过水解、羟基化、脱环氧化和葡萄糖醛酸化等反应代谢转化为毒性较低的代谢物。单端孢霉烯族毒素,例如T-2毒素和脱氧雪腐镰刀菌烯醇(DON),能迅速通过粪便和尿液排出体外。在大鼠中,给药96小时后,25%的DON经尿液排出,65%经粪便排出。 尽管绵羊摄入脱氧雪腐镰刀菌烯醇后30分钟内即可在血液中检测到,但其全身生物利用度仅为7.5%。对四只1岁雄性绵羊进行单次口服灌胃,剂量为5 mg/kg体重,并在30小时内重复采集血样。采用气相色谱-电子捕获检测器(GC-ECD)测定脱氧雪腐镰刀菌烯醇及其脱环氧代谢物。在30小时的观察期内,血浆中未检测到脱氧雪腐镰刀菌烯醇或其脱环氧代谢物。另对三只雄性绵羊进行单次静脉注射,剂量为0.5 mg/kg体重,并按照口服给药后的方法进行血液采样和分析。系统生物利用度通过浓度-时间曲线下面积与剂量的比值计算得出,口服和静脉给药均适用。血浆中检测到的去环氧代谢物占口服剂量的<0.3%,静脉给药剂量的<2%。游离脱氧雪腐镰刀菌烯醇平均占血液吸收剂量的24.8%;其余部分由去环氧代谢物或脱氧雪腐镰刀菌烯醇的葡萄糖醛酸苷结合物组成。根据两只绵羊口服5 mg/kg体重脱氧雪腐镰刀菌烯醇后36小时内收集的尿液和胆汁的回收率,脱氧雪腐镰刀菌烯醇在绵羊体内的口服吸收率约为7%。采用气相色谱-电子捕获检测器(GC/ECD)分析脱氧雪腐镰刀菌烯醇及其去环氧代谢物。尿液中回收的脱氧雪腐镰刀菌烯醇平均占给药剂量的6.9%,胆汁中回收的脱氧雪腐镰刀菌烯醇平均占给药剂量的0.11%。胆汁中仅检测到葡萄糖醛酸苷结合的脱环氧代谢物(检测限为 0.1 mg,相当于给药剂量的 0.04%)。尿液中平均回收的脱环氧代谢物或其结合物占给药剂量的 1.3%,脱氧雪腐镰刀菌烯醇或其结合物占给药剂量的 5.7%。 与绵羊和牛的低生物利用度相比,猪的生物利用度相对较高。分别在灌胃给予 0.6 mg/kg 体重的 [(14)C]脱氧雪腐镰刀菌烯醇或静脉注射 0.3 mg/kg 体重后 24 小时内收集血液、尿液、胆汁和粪便。假设放射性标记物的比例代表给药的脱氧雪腐镰刀菌烯醇的比例,并通过 GC/MS 证实了这一假设,结果显示代谢或结合反应极少。基于对三只静脉注射和三只胃内灌注动物的浓度-时间曲线下面积的测量,估计猪体内脱氧雪腐镰刀菌烯醇的平均全身生物利用度为55%。约95%的给药剂量以脱氧雪腐镰刀菌烯醇的形式回收……。 虽然尚未测定大鼠体内脱氧雪腐镰刀菌烯醇的绝对生物利用度,但96小时后,10 mg/kg体重的口服剂量中有25%在尿液中回收,表明大鼠的吸收率可能高于绵羊或牛。高效液相色谱(HPLC)和气相色谱-质谱联用(GC/MS)分析表明,0-24小时尿液中25%的放射性标记物与未代谢的脱氧雪腐镰刀菌烯醇相关,10%与脱环氧代谢物相关。同样,在96小时内,Wistar大鼠口服6 mg/kg体重的剂量后,分别有4.5%和4.4%以游离脱氧雪腐镰刀菌烯醇和脱环氧代谢物的形式从尿液中回收。 有关脱氧雪腐镰刀菌烯醇(共13种)的更多吸收、分布和排泄(完整)数据,请访问HSDB记录页面。 代谢/代谢物 在三头体重为10-15 kg的约克郡猪中,通过灌胃或腹腔注射给予脱氧雪腐镰刀菌烯醇和15-乙酰脱氧雪腐镰刀菌烯醇时,最低单次催吐剂量分别为0.050和0.075 mg/kg体重。灌胃给药后,15头猪中有3头给予15-乙酰代谢物,15头猪中有4头给予脱氧雪腐镰刀菌烯醇,所有剂量范围为20-200 μg/kg体重。腹腔注射给药后,15头猪中有9头在所有剂量下均出现呕吐。经口灌胃或腹腔注射后,脱氧雪腐镰刀菌烯醇的无观察效应剂量(NOEL)为0.025 mg/kg体重,15-乙酰代谢物的无观察效应剂量为0.050 mg/kg体重…… 脱氧雪腐镰刀菌烯醇(DON)无需活化即可产生毒性,但个体解毒能力的差异可能影响其毒性敏感性。本研究报告了对34名英国成年人尿液样本中未代谢的脱氧雪腐镰刀菌烯醇(DON)(游离DON)和DOM-1的测量结果。这些样本此前已用于分析游离DON+DON-葡萄糖醛酸苷(fD+DG)的联合浓度,浓度均>5 ng/mL。其中22名受试者连续4天采集尿液样本进行分析,而12名受试者仅采集了单次尿液样本。该子集中尿液fD+DG的平均(中位数)浓度为17.8 ng/mL(13.8 ng/mL),范围为5.0-78.2 ng/mL。在34名受试者中的23名(68%)中检测到游离DON,平均浓度为2.4 ng/mL,范围为0.5-9.3 ng/mL。在34名受试者中的1名(3%)中检测到尿液DOM-1。该个体尿液中游离DON和DG的浓度约为1%。fD+DG的总浓度与尿液中游离DON的浓度显著相关(p<0.001,R²=0.65),但与游离DON占fD+DG的百分比无显著相关性(p=0.615,R²=0.01),表明在所观察的范围内,DON的暴露水平并未影响其代谢为DG的过程。在本调查中,大多数个体尿液中未检测到DOM-1,68%的个体未能将所有摄入的DON解毒为DON-葡糖醛酸苷…… 脱氧雪腐镰刀菌烯醇-3-β-D-葡萄糖苷(D3G)是镰刀菌毒素脱氧雪腐镰刀菌烯醇(DON)的植物II期代谢产物,存在于天然污染的小麦、玉米、燕麦、大麦及其制品中。尽管D3G被认为是植物的解毒产物,但其对哺乳动物的毒性目前尚不清楚。一个主要担忧是,在哺乳动物消化过程中,D3G结合物可能水解回其有毒的前体真菌毒素DON。作者利用体外模型系统,模拟消化的不同阶段,研究了D3G在酸性条件、水解酶和肠道细菌作用下的稳定性。研究发现,D3G在37℃下至少24小时内能耐受0.2 M盐酸,表明其在哺乳动物胃中不会被水解。人胞质β-葡萄糖苷酶也无作用,而真菌纤维素酶和纤维二糖酶制剂可以裂解相当一部分D3G。更重要的是,几种乳酸菌,例如杜兰肠球菌、蒙氏肠球菌和植物乳杆菌,表现出很强的D3G水解能力。综合这些数据表明,D3G具有毒理学意义,应被视为一种隐匿性霉菌毒素。 将20% (w/w) 的大鼠盲肠内容物悬液与浓度为35 μg/mL的[(14)C]脱氧雪腐镰刀菌烯醇在厌氧条件下孵育长达24小时。采用高效液相色谱(HPLC)标准共洗脱法定量分析与脱氧雪腐镰刀菌烯醇及其脱环氧化形式相关的放射性标记物的比例。后者在加入脱氧雪腐镰刀菌烯醇后立即占所添加放射性标记物的1.3%,7小时后为29%,24小时后为90%;7小时时有60%与脱氧雪腐镰刀菌烯醇共洗脱,24小时时为2%。在另一项研究中,将1 mL浓度为1 μg/mL的脱氧雪腐镰刀菌烯醇溶液加入2 g猪大肠内容物(包括盲肠,但未具体说明)中,并在厌氧条件下培养96小时,结果表明脱氧雪腐镰刀菌烯醇并未转化为脱环氧代谢物。该研究报告称,几乎完全回收了完整的脱氧雪腐镰刀菌烯醇。而对鸡进行相同处理的肠道内容物培养96小时后,脱氧雪腐镰刀菌烯醇几乎完全转化为脱环氧代谢物。培养24小时后,在0.014 μg/mL、0.14 μg/mL和1.4 μg/mL的浓度下,转化率分别为56%、69%和70%。同样,在牛瘤胃液中孵育96小时后,35%的脱氧雪腐镰刀菌烯醇代谢为脱环氧代谢物。 有关脱氧雪腐镰刀菌烯醇(共8种代谢物)的更多代谢/代谢物(完整)数据,请访问HSDB记录页面。 生物半衰期 /猪口服后,血浆峰浓度在15-30分钟内达到,持续约9小时,然后以7.1小时的半衰期下降。 猪单次静脉注射脱氧雪腐镰刀菌烯醇(1 mg/kg体重)后,……估计消除半衰期为3.9小时。 口服25 mg/kg体重剂量的脱氧雪腐镰刀菌烯醇(DON)后,迅速被血浆吸收。最高浓度分别为 12.1 和 11.5 μg/ml,分别在 5 分钟和 15 分钟时检测到(图 1)。其清除迅速,符合双室动力学模型,分布半衰期 (t1/2α) 和清除半衰期 (t1/2β) 分别为 20.4 分钟和 11.8 小时。[5] 脱氧雪腐镰刀菌烯醇 (DON) 在肝脏(图 2A)和肾脏(图 2B)中的分布和清除动力学同样遵循双室动力学模式。与血浆中观察到的情况类似,DON 在肝脏(分别为 19.6 和 12.1 μg/g)(图 2A)和肾脏(分别为 7.6 和 9.0 μg/g)(图 2B)中的浓度在 5 至 15 分钟时达到峰值。肝脏中脱氧雪腐镰刀菌烯醇(DON)的半衰期(t1/2α)和半衰期(t1/2β)分别为22分钟和19.0小时,而肾脏中T1/2α和T1/2β分别为47分钟和20.9小时。[5] 与脾脏和肝脏类似,在上述组织中,脾脏中DON浓度最高出现在5分钟(7.3 μg/g)和15分钟(7.9 μg/g)(图3A)。T1/2α和T1/2β分别为29分钟和9.0小时。心脏中DON浓度最高出现在5分钟(6.7 μg/g)和15分钟(6.8 μg/g),T1/2α和T1/2β分别为41分钟和12.3小时(图3A)。 3B).[5] 与其他组织相比,脱氧雪腐镰刀菌烯醇 (DON) 进入大脑的速度要慢得多,且峰值浓度也低得多 (0.7-1.0 μg/g),持续时间为 5 分钟至 2 小时(图 4)。[5] 一项为期 4 周的喂养研究还评估了亚慢性膳食暴露于脱氧雪腐镰刀菌烯醇 (DON) 对血浆中该霉菌毒素浓度的影响。在以 2、5、10 和 20 mg/kg 的饲料喂养小鼠时,观察到血浆 DON 浓度呈剂量依赖性增加,浓度范围为 20 至 100 ng/ml(图 5)。所有这四个剂量均降低了体重增加(数据未显示),这与之前在 B6C3F1 小鼠中的报道一致 (Forsell 等,1986)[5] 本文提出的结果表明:脱氧雪腐镰刀菌烯醇 (DON) 在小鼠体内的吸收和分布可通过 ELISA 法进行监测。DON 暴露后迅速被组织吸收,其最大浓度依次为:肝脏 > 血浆 > 肾脏 > 脾脏 > 心脏 > 脑。除脑组织外,DON 在所有组织中的初始清除速度均较快,随后清除速度相对较慢。本文数据与我们实验室之前在小鼠中进行的 [3H]-DON 研究(Azcona-Olivera 等,1995)高度一致。该研究也报道了 DON 的双室动力学特征,即在暴露于 5 mg/kg 体重(t1/2α = 21.6 分钟,t1/2β = 7.6 小时)和 25 mg/kg 体重(t1/2α = 21.6 分钟,t1/2β = 7.6 小时)的 DON 后,血浆中 DON 的初始清除速度较快,而最终清除速度较慢。分别为 33.6 分钟和 88.9 小时)。当将本研究中前 4 小时内血浆、肝脏和肾脏中 DON 的吸收和清除情况与上述 25 mg/kg 体重剂量下的研究结果进行图表比较时,发现两者非常相似(图 6)。因此,ELISA 法与放射性标记法在监测小鼠体内 DON 组织浓度和毒代动力学方面具有可比性。该免疫测定方法的主要优点是:(1) 灵敏度和特异性很高;(2) 采用简便的样品制备方案,无需复杂的净化步骤;(3) 无需蒸发或浓缩。[5] 小鼠单次口服 25 mg/kg 体重剂量后,脱氧雪腐镰刀菌烯醇 (DON) 被迅速吸收和分布。血浆中 DON 浓度最高 (12 μg/mL) 在 5 至 15 分钟内达到。[5] 血浆清除率随后药物动力学符合双室模型,快速分布半衰期 (t₁/₂α) 为 20.4 分钟,较慢的末端消除半衰期 (t₁/₂β) 为 11.8 小时。给药后 8 小时和 24 小时,血浆中药物浓度分别剩余最大浓度的 5% 和 2%。[5] 在给药后 5 分钟至 24 小时内,血浆、肝脏、脾脏和脑组织中均可检测到脱氧雪腐镰刀菌烯醇 (DON);在给药后 5 分钟至 8 小时内,心脏和肾脏中均可检测到 DON。[5] 给药后 5 分钟,各组织浓度(平均值 ± 标准误)分别为:肝脏 19.5 ± 1.9 μg/g,肾脏 7.6 ± 0.5 μg/g,脾脏 7.3 ± 0.8 μg/g,心脏 6.8 ± 0.9 μg/g,脑组织 0.8 ± 0.1 μg/g。分布和清除动力学组织中的DON水平与血浆中的DON水平相似。[5] 在亚慢性喂养研究中,血浆DON水平随日粮中DON浓度(0至20 mg/kg日粮)的增加呈剂量依赖性增加。[5] |
| 毒性/毒理 (Toxicokinetics/TK) |
相互作用
脱氧雪腐镰刀菌烯醇 (DON) 和伏马菌素 (FB) 是镰刀菌属产生的最常见的真菌毒素,也是动物饲料中最常见的共存物质。本研究考察了这些真菌毒素对仔猪的毒性,并分析了包括血浆生化指标、器官组织病理学和免疫反应在内的多个参数。24 头 5 周龄仔猪被随机分为四组,分别饲喂不同的饲料,持续 5 周:对照组、分别添加 DON (3 mg/kg)、FB (6 mg/kg) 或两种毒素的混合饲料组。在试验的第 4 天和第 16 天,对仔猪进行卵清蛋白皮下免疫,以评估其特异性免疫反应。不同的饲料对仔猪的生长性能没有影响,对血液学和生化指标的影响也很小。然而,DON 和 FB 均在仔猪的肝脏、肺脏和肾脏中诱发了组织病理学损伤。当两种霉菌毒素同时存在时,肝脏受到的影响更为显著。受污染的饲料还会改变疫苗接种后的特异性免疫反应,表现为血浆中抗卵清蛋白IgG水平降低以及抗原刺激后淋巴细胞增殖减少。由于细胞因子在免疫中发挥关键作用,我们在实验结束时通过RT-PCR检测了IL-8、IL-1β、IL-6和巨噬细胞炎症蛋白-1β的表达水平。结果显示,在暴露于多种污染饲料的仔猪脾脏中,这四种细胞因子的表达显著降低。综上所述,我们的数据表明,与摄入单一污染饲料相比,摄入多种污染饲料会导致更严重的组织病理学损伤和更高的免疫抑制。 博维菌素(BEA)、脱氧雪腐镰刀菌烯醇(DON)和T-2毒素(T-2)是重要的食源性霉菌毒素,与人类健康密切相关。本研究采用四唑盐(MTT)法和中性红(NR)法,在永生化仓鼠卵巢细胞(CHO-K1)中检测了单一及联合真菌毒素(BEA、DON 和 T-2)在暴露 24、48 和 72 小时后的急性毒性。MTT 法和 NR 法测得所有真菌毒素的 IC50 值分别为 0.017~12.08 μM 和 0.042~17.22 μM。单一及联合真菌毒素均表现出对 CHO-K1 细胞的显著剂量依赖性细胞毒性作用。当单独检测霉菌毒素时,T-2 在两个测试终点均表现出最强的 IC50 值(0.017 至 0.052 μM),其次是 DON(0.53 至 2.30 μM),而 BEA 的 IC50 值最弱(2.77 至 17.22 μM)。另一方面,采用等效线图法评估了细胞毒性相互作用。在急性二元试验中,DON+BEA(CI=1.60-25.07)和 DON+T-2(CI=1.74-7.71)在暴露 24、48 和 72 小时后均表现出拮抗作用。相比之下,BEA+T-2 二元组合(CI=0.35-0.78)在所有测试暴露时间点均表现出协同作用。三元化合物 BEA+DON+T-2 组合在暴露 24 小时和 48 小时后表现出协同效应(CI=0.47-0.86);然而,在较低剂量下暴露 72 小时后则表现出中等程度的拮抗效应(CI=1.14-1.60)。这些结果提供了定量证据,表明 BEA、DON 和 T-2 之间可能存在重要的相互作用,且这种相互作用取决于暴露时间。组合指数-等效线图方程法可作为食品风险评估的有效工具。由于 BEA、DON 和 T-2 具有强烈的毒性作用,从真菌毒理学的角度来看,它们的联合暴露可能是人类多种疾病发生的重要诱因,尤其是在长期暴露之后。 本研究在 3 项试验中,以体重 15-18 克的 21 日龄雄性 ICR 小鼠为对象,研究了赛庚啶(一种 5-羟色胺 2 受体拮抗剂和已知的食欲刺激剂)减轻脱氧雪腐镰刀菌烯醇不良反应的能力。 10只小鼠一组,饲喂含有赛庚啶和脱氧雪腐镰刀菌烯醇(纯度99%)组合的饲料,脱氧雪腐镰刀菌烯醇的剂量为4-16 mg/kg(相当于0.6-2.4 mg/kg体重/天),赛庚啶的剂量为1.2-20 mg/kg(相当于0.19-3 mg/kg体重)。在添加脱氧雪腐镰刀菌烯醇前2天,先在饲料中添加赛庚啶,然后两种物质同时添加12天。赛庚啶能有效抵消脱氧雪腐镰刀菌烯醇引起的采食量减少,但仅在特定剂量下有效。当饲料中脱氧雪腐镰刀菌烯醇的剂量为4 mg/kg时,赛庚啶的最佳剂量为1.2-2.5 mg/kg饲料。当饲料中脱氧雪腐镰刀菌烯醇(DON)浓度为 8 mg/kg 时,所需剂量为 2.5 mg/kg;当饲料中 DON 浓度为 12 mg/kg 时,所需剂量为 2.5-5.0 mg/kg;当饲料中 DON 浓度为 16 mg/kg 时,所需剂量为 5-10 mg/kg。在较低剂量的赛庚啶(5 mg/kg)下,无论单独使用还是与最低测试剂量的 DON 联合使用,均可观察到体重略有增加,但在较高浓度的 DON 下则未观察到此现象。由此得出结论:血清素能机制可能介导了 DON 引起的采食量减少。研究发现赛庚啶能显著减弱脱氧雪腐镰刀菌烯醇的作用,表明5-羟色胺2受体参与了这一过程。 本研究使用3-6头体重60公斤的猪,研究饲料中单独或联合添加纯化的脱氧雪腐镰刀菌烯醇和赭曲霉毒素A对猪的健康影响,以及摄入后90天的残留情况。猪分别饲喂以下饲料:含0.1毫克/千克赭曲霉毒素A和1毫克/千克脱氧雪腐镰刀菌烯醇的饲料(分别相当于每公斤体重0.004毫克赭曲霉毒素A和0.04毫克脱氧雪腐镰刀菌烯醇);单独饲喂0.1毫克/千克赭曲霉毒素A的饲料;或单独饲喂1毫克/千克脱氧雪腐镰刀菌烯醇的饲料。另设两个对照组,饲喂不含赭曲霉毒素A和脱氧雪腐镰刀菌烯醇的饲料。饲料中添加霉菌毒素的猪未出现临床或血液学变化。同时添加两种霉菌毒素的猪出现胃黏膜充血,且每个处理组中均有一头猪出现肾小管上皮细胞改变。虽然病理损伤较少,但委员会指出,本研究的动物数量较少。观察到的伪狂犬病(奥耶斯基病或“狂犬病”)抗体滴度(作为免疫系统影响的指标)表明,非特异性防御机制未受影响。同时接受两种毒素处理的动物肾脏中赭曲霉毒素A的平均浓度比单独接受赭曲霉毒素A处理的组高约50%,表明可能存在相互作用。在同时接受两种霉菌毒素处理的动物的肌肉中,赭曲霉毒素A的浓度似乎也略有升高。 有关脱氧雪腐镰刀菌烯醇(DEOXYNIVALENOL)的更多相互作用(完整)数据(共6项),请访问HSDB记录页面。 非人类毒性值 小鼠(雄性)腹腔注射LD50:70 mg/kg 小鼠(雌性)腹腔注射LD50:76.7 mg/kg 小鼠口服LD50:46 mg/kg 小鼠皮下注射LD50:45 mg/kg 有关脱氧雪腐镰刀菌烯醇(DEOXYNIVALENOL)的更多非人类毒性值(完整)数据(共11项),请访问HSDB记录页面。 本研究中使用的25 mg/kg体重的急性口服剂量相当于先前报道的B6C3F1雌性小鼠LD₅₀的三分之一。 [5] 已知DON可引起厌食、肠胃炎、呕吐和血液系统疾病。[5] |
| 参考文献 |
|
| 其他信息 |
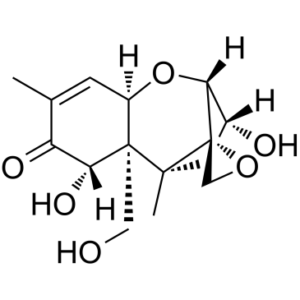
脱氧雪腐镰刀菌烯醇(DON)是一种由镰刀菌属真菌产生的单端孢霉烯族毒素,小麦、大麦、玉米及其制品易受其污染。它是一种真菌毒素。DON是一种单端孢霉烯族化合物,属于环状酮、仲α-羟基酮、伯醇、烯酮和三醇类化合物。
已有报道称,在禾谷镰刀菌(Fusarium graminearum)、腐霉镰刀菌(Fusarium culmorum)和眼虫(Euglena gracilis)中均检测到了DON,并有相关数据可供参考。 作用机制 本研究旨在探讨用脱氧雪腐镰刀菌烯醇(DON)处理的胸腺瘤细胞与未处理(对照)细胞相比,其蛋白质组学分析是否能揭示差异表达的蛋白质,从而有助于更好地理解其毒性机制。为此,将小鼠胸腺瘤细胞系EL4暴露于0.5 μM DON中6小时。EL4细胞暴露于DON后,共有30种蛋白质受到影响。这些蛋白质大多表达上调,包括关键代谢酶(例如脂肪酸合成酶、醛糖还原酶、氨甲酰磷酸合成酶、葡萄糖-6-磷酸异构酶)、分子伴侣(例如HSP9AB1和HSP70)、参与蛋白质折叠的酶(PDI和ERO1-1α)以及参与蛋白质降解的蛋白质(泛素结合酶(E1)和蛋白酶体α亚基1型)。此外,还发现分子量为60 kDa的IgE结合蛋白和转录因子My结合蛋白1a(MYBBP1A)的表达也受到DON的上调。观察到MYBBP1A(一种已知的多种转录因子抑制剂,例如PGC-1α、C-myb和NF-κB家族的p65)的表达上调,提示该蛋白可能在DON毒性机制中发挥作用。 脱氧雪腐镰刀菌烯醇(DON)是谷物中最常见的单端孢霉烯族毒素之一,与人类和家畜的真菌毒素中毒有关。低剂量毒性表现为体重增长减少、营养利用率降低和免疫功能受损。人类食品中DON污染的程度和模式表明,DON的摄入构成了一个公共卫生问题。DON在加工和烹饪过程中的稳定性解释了其在人类食品中的大量存在。/作者/在此通过证明该毒素在口服和中枢给药后均会影响摄食行为、体温和运动活性,从而表征了DON中毒。利用 c-Fos 表达谱分析,作者鉴定了 DON 激活的神经元结构,并观察到毒素激活的神经元群模式与炎症信号诱导的模式相似。通过实时 PCR,作者报告了 DON 诱导中枢炎症的证据,表现为白细胞介素-1β、白细胞介素-6、肿瘤坏死因子-α、环氧合酶-2 和微粒体前列腺素合成酶-1 (mPGES-1) 信使 RNA 的显著上调。然而,使用对细胞因子诱导的疾病行为具有抵抗力的 mPGES-1 基因敲除小鼠沉默前列腺素 E2 信号通路,并未改变对该毒素的反应。这些结果表明,尽管存在诸多相似之处,但DON中毒后观察到的行为变化与炎症细胞因子诱发的经典疾病行为有所不同。 单端孢霉烯族化合物对活跃分裂的细胞(例如肠隐窝上皮细胞和造血细胞)具有毒性。其细胞毒性与化合物与真核细胞核糖体结合导致蛋白质合成受损,或细胞膜功能障碍有关。蛋白质合成抑制与免疫细胞中不稳定蛋白和调节蛋白(例如IL-2)的诱导有关。极低浓度的单端孢霉烯族化合物即可损害细胞膜上小分子的转运。/单端孢霉烯族化合物/ 大多数单端孢霉烯族化合物抑制蛋白质合成,其效力取决于结构取代基,并且需要C9-C10位上的不饱和键以及12,13-环氧环的完整性。单端孢霉烯族毒素与真核生物核糖体的60S亚基结合,干扰肽基转移酶的活性。脱氧雪腐镰刀菌烯醇(DON)在C-4位缺乏取代基,可抑制肽链延伸。抑制蛋白质合成被认为是包括DON在内的单端孢霉烯族毒素的主要毒性作用。DON抑制兔网织红细胞蛋白质合成的ID50为2 μg/mL。体外实验表明,DON的毒性比T-2毒素低约100倍,而T-2毒素因其对大分子的影响而得到更广泛的研究。由于亲脂性及其他可能的影响,脱氧雪腐镰刀菌烯醇在体内的毒性大于其在体外对蛋白质合成的影响所预期的毒性。 有关脱氧雪腐镰刀菌烯醇(共10种)的更多作用机制(完整)数据,请访问HSDB记录页面。 脱氧雪腐镰刀菌烯醇(DON,呕吐毒素)是一种单端孢霉烯族真菌毒素,通常由禾谷镰刀菌产生,存在于受污染的谷物中,例如小麦、大麦和玉米。[5] 其在食品中的存在是全球范围内一个重要的人类健康问题。[5] 所有动物物种均对DON敏感,其敏感性排序为:猪 > 小鼠 > 大鼠 > 家禽 > 反刍动物。代谢、吸收、分布和排泄方面的差异可能是造成这种差异的原因。 [5] 本研究描述的竞争性直接ELISA方法是一种灵敏且特异的检测生物样品中DON的工具,在添加DON的组织样品中回收率可达90%或更高。[5] |
| 分子式 |
C15H20O6
|
|---|---|
| 分子量 |
296.3157
|
| 精确质量 |
296.125
|
| 元素分析 |
C, 60.80; H, 6.80; O, 32.40
|
| CAS号 |
51481-10-8
|
| 相关CAS号 |
Deoxynivalenol-13C15;911392-36-4
|
| PubChem CID |
40024
|
| 外观&性状 |
Fine needles from ethyl acetate + petroleum ether
Crystals from methanol (aqueous) |
| 密度 |
1.5±0.1 g/cm3
|
| 沸点 |
543.9±50.0 °C at 760 mmHg
|
| 熔点 |
151-153ºC
|
| 闪点 |
206.9±23.6 °C
|
| 蒸汽压 |
0.0±3.3 mmHg at 25°C
|
| 折射率 |
1.632
|
| 来源 |
Fusarium specie; Fusarium graminearum, Fusarium culmorum, and Euglena gracilis
|
| LogP |
-1.41
|
| tPSA |
99.52
|
| 氢键供体(HBD)数目 |
3
|
| 氢键受体(HBA)数目 |
6
|
| 可旋转键数目(RBC) |
1
|
| 重原子数目 |
21
|
| 分子复杂度/Complexity |
558
|
| 定义原子立体中心数目 |
7
|
| SMILES |
O1C([H])([H])[C@]21[C@@]1([H])[C@@]([H])(C([H])([H])[C@]2(C([H])([H])[H])[C@@]2(C([H])([H])O[H])[C@@]([H])(C(C(C([H])([H])[H])=C([H])[C@@]2([H])O1)=O)O[H])O[H]
|
| InChi Key |
LINOMUASTDIRTM-QGRHZQQGSA-N
|
| InChi Code |
InChI=1S/C15H20O6/c1-7-3-9-14(5-16,11(19)10(7)18)13(2)4-8(17)12(21-9)15(13)6-20-15/h3,8-9,11-12,16-17,19H,4-6H2,1-2H3/t8-,9-,11-,12-,13-,14-,15+/m1/s1
|
| 化学名 |
(3alpha,7alpha)-3,7,15-Trihydroxy-12,13-epoxytrichothec-9-en-8-one
|
| 别名 |
Vomitoxin; 3-epi-DON; DEOXYNIVALENOL; 51481-10-8; 4-Deoxynivalenol; CRIS 7801; Dehydronivalenol
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
Ethanol : ~30 mg/mL (~101.24 mM)
DMSO : ~25 mg/mL (~84.37 mM) |
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.5 mg/mL (8.44 mM) (饱和度未知) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 25.0 mg/mL澄清DMSO储备液加入到400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: ≥ 2.5 mg/mL (8.44 mM) (饱和度未知) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 例如,若需制备1 mL的工作液,可将 100 μL 25.0 mg/mL澄清DMSO储备液加入900 μL 20% SBE-β-CD生理盐水溶液中,混匀。 *20% SBE-β-CD 生理盐水溶液的制备(4°C,1 周):将 2 g SBE-β-CD 溶解于 10 mL 生理盐水中,得到澄清溶液。 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 3.3747 mL | 16.8737 mL | 33.7473 mL | |
| 5 mM | 0.6749 mL | 3.3747 mL | 6.7495 mL | |
| 10 mM | 0.3375 mL | 1.6874 mL | 3.3747 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1
































 COA
COA
 463611831
463611831
