| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| 500mg |
|
||
| Other Sizes |
|
| 靶点 |
G-quadruplexe (Kd = 490 nM)
|
|---|---|
| 体外研究 (In Vitro) |
Pyridostatin (RR82) 盐酸盐(10 μM;48 小时)可诱导细胞周期停滞[1]。
Pyridostatin 盐酸盐是一种小分子,可选择性结合 G-四链体 DNA,形成稳定 G-四链体结构的复合物。盐酸吡啶他汀会引起神经突回缩、突触丧失和剂量依赖性神经元死亡。 PyridostatinHCl 可诱导培养的原代神经元中 DNA 双链断裂的形成。保护和固定神经元基因组的 BRCA1 蛋白在转录水平上被盐酸吡咯抑素(1–5 μM,过夜)显着下调[3]。 |
| 体内研究 (In Vivo) |
Pyridostatin对BRCA2缺陷异种移植物具有抗肿瘤活性[4]
结合和稳定G4s的化合物已被证明对小鼠建立的BRCA1/2缺失异种移植物肿瘤有活性(RHPS4和CX‐5461)。然而,这些尚未被证明对BRCA突变患者有益。此外,BRCA突变的肿瘤很难治疗,因为它们对靶向治疗(例如PARP抑制剂;PARPi)。因此,必须找到新的G4配体,不仅可以消除BRCA缺陷肿瘤,还可以抵抗耐药疾病。我们之前发表的结果(Zimmer et al, 2016)表明,G4配体pyridostatin对体外BRCA2缺陷细胞具有特异性毒性。在这项研究中,研究人员评估了pyridostatin在体内消除BRCA2缺陷异种移植物肿瘤中的潜力。为了解决这个问题,我们使用等基因BRCA2 +/+ (BRCA2‐精通)和BRCA2−/−(BRCA2‐缺乏)人结直肠癌DLD1细胞在CB17‐SCID小鼠中产生异种移植物(图1A和B)。研究人员广泛优化了pyridostatin在体内使用的条件,并确定了7.5 mg/kg/天的剂量计划静脉注射连续5天,然后休息2天,第二个5天的治疗耐受性良好。没有明显的体重减轻,没有不良的临床症状(附录表S1)。在这些条件下,研究人员发现pyridostatin有效且特异性地抑制BRCA2‐缺陷DLD1细胞建立的异种移植物肿瘤的生长(图1B)。作为对照,研究人员使用了PARPi talazoparib,该药物以其根除小鼠BRCA1/2缺失肿瘤的能力而闻名(Shen et al ., 2013),最近被许可用于携带BRCA1/2种系突变的转移性乳腺癌患者(Litton et al ., 2018)。pyridostatin对BRCA2 -缺陷肿瘤的抗肿瘤作用与talazoparib相似,两种药物都不会损害BRCA2 -精通肿瘤的生长。 此外,研究人员使用第二种肿瘤模型研究了pyridostatin在体内的反应,该模型是由等基因BRCA2 +/+和BRCA2 - / -结直肠癌HCT116细胞建立的(Xu et al ., 2014)。Pyridostatin显示出对BRCA2缺失HCT116细胞源性肿瘤的选择性毒性(附录图S1A和B;附录表S2),其作用与DLD1细胞来源的异种移植物相似[4] 研究人员之前的研究表明,pyridostatin治疗会导致HR修复受损的细胞(包括BRCA2缺陷细胞)中DNA损伤的积累(Zimmer等,2016)。一致地,免疫组织化学(IHC)分析显示,BRCA2‐缺乏,但不是BRCA2‐完全,肿瘤在暴露于pyridostatin或talazoparib时显示出DNA损伤标记γH2AX水平增加(附录图S1C-F)。这些结果表明,pyridostatin不仅可以特异性抑制细胞的生长(Zimmer等,2016),还可以特异性抑制缺乏BRCA2的肿瘤,并通过造成DNA损伤在体内起作用[4]。 |
| 细胞实验 |
细胞系: 超过 60 种不同的癌细胞系
浓度: 10 μM 孵育时间: 48 小时 结果: 主要在超过 60 种不同的癌细胞系的细胞周期的 G2 期积累。 |
| 动物实验 |
CB17-SCID 小鼠
7.5 mg/kg i.v. 体内异种移植实验[4] CB17-SCID 小鼠(CB17/Icr-Prkdcscid/IcrIcoCrl,雄性或雌性)和 FVB 雌性小鼠购自 Charles River Laboratories。小鼠饲养于高效空气微粒过滤器 (HEPA) 过滤的笼架中,并喂以高压灭菌的实验室啮齿动物饲料。[4] 为了构建源自 DLD1 和 HCT116 BRCA2 功能正常或缺陷细胞的异种移植模型,将 5 × 10⁶ 个细胞/只注射到 6 周龄 CB17-SCID 雄性小鼠的后腿肌肉中。当肿瘤体积达到约 250 mm³ 时,将小鼠随机分组开始治疗。[4] 为了构建 PARP 抑制剂耐药小鼠肿瘤模型,将 4 × 10⁶ 个 KP3.33 (Brca1 +/+) 细胞或 KB1PM5 (Brca1 −/− Tp53bp1 −/−) 小鼠乳腺肿瘤细胞肌内注射到 6 周龄 FVB 雌性小鼠的后肢肌肉中。每个实验组包含 5 只小鼠。当肿瘤体积达到约 250 mm³ 时,将小鼠随机分组并开始治疗。[4] 为了构建 MDA-MB-436 细胞来源的异种移植瘤,将 4 × 10⁶ 个细胞肌内注射到 6 周龄 CB17-SCID 雌性小鼠的后肢肌肉中。当肿瘤体积达到约 220 mm³ 时(细胞注射后 6 天),开始治疗。每个实验组包含五只小鼠。[4] Talazoparib(BMN 673,Selleckchem)溶于10%二甲基乙酰胺、6% HS溶液和84% PBS中,以0.33 mg/kg/天的剂量口服给药,连续给药五天,然后停药两天,再给药五天(Wang et al, 2016)。吡啶斯他汀溶于生理盐水中,以7.5 mg/kg/天的剂量静脉注射给药,连续给药五天,然后停药两天,再给药五天。 NU-7441(Selleckchem)溶于5% DMSO、40% PEG300和5% Tween-80的混合溶液中,以10 mg/kg/天的剂量腹腔注射,连续给药5天,之后停药2天,再继续给药5天(Zhao等,2006)。紫杉醇溶于生理盐水中,于治疗的第1天和第8天以20 mg/kg/天的剂量静脉注射(Bizzaro等,2018)。与其他化合物联合用药时,紫杉醇于治疗的第5天和第12天静脉注射,吡啶斯他汀和NU-7441分别静脉注射和腹腔注射,连续给药4天,之后停药3天,再继续给药4天。 NU-7441 在给予吡啶斯他汀前 2 小时给药。在指定时间点,使用游标卡尺测量肿瘤的二维体积,并根据肿瘤体积估算肿瘤重量(1 mg = 1 mm³)。采用学生 t 检验(非配对,双尾)进行单组两两比较。当 P < 0.05 时,差异被认为具有统计学意义。小鼠的生存曲线采用 Kaplan-Meier 法绘制,并使用对数秩检验评估统计学意义。数据采用 GraphPad Prism 8.3 软件绘制。[4] PDTX 模型构建[4] 前瞻性地收集了携带 gBRCA 基因的乳腺癌患者的新鲜肿瘤样本,用于在机构伦理审查委员会 (IRB) 批准的方案和相关知情同意书下,或由剑桥郡国家研究伦理服务中心 (REC) 批准(REC 参考编号:08/H0308/178)(Bruna 等,2016)移植到小鼠体内。VHI0179 患者来源肿瘤异种移植模型 (PDTX) 由携带 BRCA1 种系截断且因 REV7 突变而对奥拉帕尼耐药的患者乳腺肿瘤构建而成。所有患者均签署了书面知情同意书,实验符合世界医学协会《赫尔辛基宣言》和美国卫生与公众服务部《贝尔蒙特报告》中规定的原则。将冷冻肿瘤组织块(15–20 mm³)包被Matrigel基质胶后,通过腰背部一侧皮下切口植入一只6周龄CB17-SCID雌性小鼠体内。当肿瘤体积达到约400 mm³时,将肿瘤从处死的小鼠体内取出,切成约15–20 mm³的组织块,再次皮下植入14只CB17-SCID雌性小鼠体内。当肿瘤体积达到约200 mm³时,将小鼠随机分为载体组和治疗组,开始治疗。每个实验组包含7只小鼠。 |
| 参考文献 |
|
| 其他信息 |
能够稳定端粒DNA G-四链体形成的配体具有作为癌症治疗药物的潜力,因为端粒酶(一种在许多癌细胞中过度表达的酶)无法延伸G-四链体结构。因此,了解小分子与这些结构结合的动力学、热力学和力学性质至关重要,但传统的整体分析方法无法同时测量这些性质。本文中,我们采用激光镊子技术研究了此类相互作用。通过力跃变法,我们观察到吡啶斯他汀促进了端粒G-四链体的折叠。吡啶斯他汀结合的G-四链体机械稳定性增强,使得我们能够测定其解离常数K(d)为490 ± 80 nM。通过类Hess过程获得的结合自由能变化表明,吡啶斯他汀的K(d)与上述结果相同,而较弱的配体RR110的K(d)为42 ± 3 µM。我们预期,这一单分子平台能够提供配体结合的生物大分子的力学、动力学和热力学性质的详细信息,这些性质具有生物学意义。[1]
富含鸟嘌呤的DNA序列能够在体外形成非沃森-克里克碱基结构,这些序列在人类基因组中普遍存在。然而,此类结构是否正常存在于哺乳动物细胞中,几十年来一直是研究的热点。本文表明,G-四链体相互作用药物吡啶斯他汀通过诱导复制和转录依赖性DNA损伤,促进人类癌细胞的生长停滞。对DNA损伤标记物γH2AX的染色质免疫沉淀测序分析,揭示了吡啶斯他汀诱导的损伤位点在基因组范围内的分布,并表明吡啶斯他汀靶向含有易于形成G-四链体序列簇的基因体。因此,吡啶斯他汀调节了这些基因的表达,包括原癌基因SRC。我们观察到吡啶斯他汀降低了人乳腺癌细胞中SRC蛋白的丰度和SRC依赖的细胞运动能力,证实了SRC是该药物的作用靶点。我们采用无偏倚的方法来确定药物的基因组作用位点,为发现功能性DNA-药物相互作用建立了框架。[2] 能够建立潜伏感染的病毒已经进化出独特的机制来逃避宿主的免疫识别。这些病毒的维持蛋白会将其合成水平调节到足以维持持续感染但又低于宿主免疫检测阈值的水平。目前尚不清楚控制这种病毒潜伏期精细调控的机制。本文中,我们发现编码γ疱疹病毒维持蛋白的mRNA在其开放阅读框内包含簇状的特殊结构元件——G-四链体,这些G-四链体负责顺式调控病毒mRNA的翻译。通过研究Epstein-Barr病毒编码的核抗原1 (EBNA1) mRNA,我们发现使用反义寡核苷酸破坏G-四链体可增加EBNA1 mRNA的翻译。相反,用G-四链体稳定剂小分子吡啶斯他汀预处理则可降低EBNA1的合成,这凸显了病毒编码转录本中G-四链体作为翻译调控和免疫逃逸的独特调控信号的重要性。此外,这些发现提示了以病毒开放阅读框(ORF)内RNA结构为靶点的替代治疗策略。[3] BRCA1或BRCA2 (BRCA1/2)功能受损的细胞会积累停滞的复制叉,导致复制相关的DNA损伤和基因组不稳定,这是BRCA1/2突变肿瘤的特征。针对BRCA1/2突变肿瘤的靶向治疗正是利用了这种脆弱性,通过引入额外的DNA损伤来发挥作用。由于BRCA1或BRCA2缺失会导致同源重组(HR)修复受损,这些损伤对肿瘤细胞具有特异性致死性,但对健康组织无害。近年来,结合并稳定G-四链体(G4)的配体作为一类化合物脱颖而出,它们能够选择性地清除缺乏BRCA1或BRCA2的细胞和肿瘤。吡啶斯他汀是一种能够结合G4的小分子,在体外对BRCA1/2缺陷细胞具有特异性毒性。然而,其体内活性尚未得到评估。本文中,我们证实吡啶斯他汀对BRCA1/2缺陷肿瘤,包括已获得PARP抑制剂(PARPi)耐药性的患者来源异种移植瘤,均表现出高度特异性活性。从机制上讲,我们证实吡啶斯他汀会干扰DNA复制,导致DNA双链断裂(DSB),而这些DSB在BRCA1/2缺失的情况下可通过经典的非同源末端连接(C-NHEJ)途径进行修复。与此一致的是,DNA-PKcs(C-NHEJ激酶活性的核心组分)的化学抑制剂与吡啶斯他汀具有协同作用,可清除BRCA1/2缺陷细胞和肿瘤。此外,我们还证实,当BRCA1或BRCA2被敲除时,吡啶斯他汀会触发cGAS/STING依赖的先天免疫反应。紫杉醇是一种常用的癌症化疗药物,可增强吡啶斯他汀的体内毒性。总的来说,我们的研究结果表明,吡啶斯他汀是一种适合进一步开发治疗的化合物,可以单独使用,也可以与紫杉醇和 DNA-PKcs 抑制剂联合使用,以造福携带 BRCA1/2 突变的癌症患者。[4] |
| 分子式 |
C31H37CL5N8O5
|
|---|---|
| 分子量 |
778.9411
|
| 精确质量 |
778.13
|
| CAS号 |
1781882-65-2
|
| 相关CAS号 |
Pyridostatin;1085412-37-8;Pyridostatin TFA;1472611-44-1
|
| PubChem CID |
78243739
|
| 外观&性状 |
White to off-white solid
|
| tPSA |
203
|
| 氢键供体(HBD)数目 |
10
|
| 氢键受体(HBA)数目 |
11
|
| 可旋转键数目(RBC) |
13
|
| 重原子数目 |
49
|
| 分子复杂度/Complexity |
850
|
| 定义原子立体中心数目 |
0
|
| SMILES |
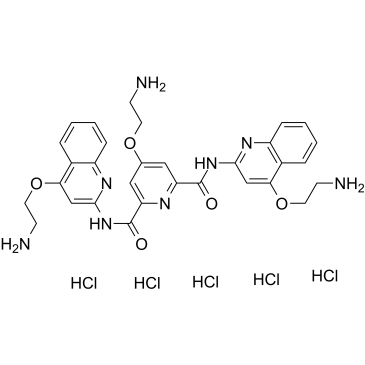
C1=CC=C2C(=C1)C(=CC(=N2)NC(=O)C3=CC(=CC(=N3)C(=O)NC4=NC5=CC=CC=C5C(=C4)OCCN)OCCN)OCCN.Cl.Cl.Cl.Cl.Cl
|
| InChi Key |
SRIZPFGTXSQRFM-UHFFFAOYSA-N
|
| InChi Code |
InChI=1S/C31H32N8O5.5ClH/c32-9-12-42-19-15-24(30(40)38-28-17-26(43-13-10-33)20-5-1-3-7-22(20)36-28)35-25(16-19)31(41)39-29-18-27(44-14-11-34)21-6-2-4-8-23(21)37-29;;;;;/h1-8,15-18H,9-14,32-34H2,(H,36,38,40)(H,37,39,41);5*1H
|
| 化学名 |
4-(2-aminoethoxy)-2-N,6-N-bis[4-(2-aminoethoxy)quinolin-2-yl]pyridine-2,6-dicarboxamide;pentahydrochloride
|
| 别名 |
RR82 hydrochloride; Pyridostatin hydrochloride; 1781882-65-2; Pyridostatin (hydrochloride); RR82 hydrochloride; 4-(2-aminoethoxy)-2-N,6-N-bis[4-(2-aminoethoxy)quinolin-2-yl]pyridine-2,6-dicarboxamide;pentahydrochloride; Pyridostatin pentahydrochloride; RR-82 hydrochloride; 4-(2-aminoethoxy)-N2,N6-bis[4-(2-aminoethoxy)quinolin-2-yl]pyridine-2,6-dicarboxamide pentahydrochloride
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month 注意: 请将本产品存放在密封且受保护的环境中(例如氮气保护),避免吸湿/受潮。 |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
H2O: ~50 mg/mL (~64.2 mM)
|
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: 100 mg/mL (128.38 mM) in PBS (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液; 超声助溶。
请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 1.2838 mL | 6.4190 mL | 12.8380 mL | |
| 5 mM | 0.2568 mL | 1.2838 mL | 2.5676 mL | |
| 10 mM | 0.1284 mL | 0.6419 mL | 1.2838 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
|
|
|
|
|
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1





































 463611831
463611831
