| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 5mg |
|
||
| 10mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| Other Sizes |
|

| 靶点 |
CYP2C8 (Ki = 4.82 μM); CYP2J2 (Ki = 5.75 μM)
|
|---|---|
| 体外研究 (In Vitro) |
Salvianolic Acid C 对 CYP2C8 和 CYP2J2 的 Ki 值分别为 4.82 和 5.75 μM,使其成为 CYP2C8 的非竞争性校正剂和 CYP2J2 的中度混合校正剂 [1]。 5 μM 丹酚酸 C 大大减少了 LPS 产生的 NO 的产生。丹酚酸 C 显着降低了 iNOS 的表达。丹酚酸 C 可防止 LPS 引起的 TNF-α、IL-1β、IL-6 和 IL-10 的过度产生。 LPS 产生的 NF-κB 活化被丹酚酸 C 抑制。在 BV2 小行星细胞中,C 还可以上调 HO-1 和 Nrf2 的表达 [2]。
丹酚酸C/SalC对细胞活力的影响[2] MTT法检测SalC对BV2小胶质细胞活力的影响。结果表明,1 μg/mL LPS对BV2小胶质细胞活力无明显影响。在0 ~ 20 μM浓度范围内,SalC对细胞活力没有影响(图1B),而在50和100 μM浓度范围内,SalC略微降低细胞活力。 SalC/丹酚酸C对lps处理BV2细胞NO、iNOS、PGE2和COX-2水平的影响[2] 采用Griess试剂测定NO释放量。如图2A所示,LPS刺激BV2细胞上清液中NO含量较对照组明显上调。1和5 μM SalC能显著抑制LPS诱导的NO生成(P < 0.01,P < 0.01)。为了明确抑制NO产生的原因,我们采用免疫荧光和Western方法进一步研究了iNOS的表达。如图2B-D所示,正常对照组iNOS表达极低,但LPS刺激可显著上调iNOS表达。SalC显著降低iNOS的表达。以上结果表明,SalC对NO生成的抑制作用是通过抑制iNOS表达实现的。进一步研究了SalC对PGE2和COX-2的影响。ELISA法检测PGE2的分泌。如图2E所示,LPS刺激导致细胞中PGE2含量显著增加(P < 0.01)。1和5 μM SalC显著降低lps诱导的PGE2上调(P < 0.05,P < 0.01)。Western blot法检测COX-2的表达。SalC显著降低LPS刺激诱导的COX-2表达(图2F)。以上结果表明,SalC可以显著抑制iNOS和COX-2的表达,从而阻断NO和PGE2的合成。 SalC/丹酚酸C抑制lps诱导的TNF - α、IL - 1β、IL - 6和IL - 10过量生成[2] 然后测量BV2小胶质细胞中TNF - α、IL - 1β、IL - 6和IL - 10的产生。结果显示,LPS刺激可显著增加BV2小胶质细胞中TNF - α、IL - 1β、IL - 6和IL - 10的产生(P < 0.01,图3A-D),而SalC可显著降低BV2小胶质细胞中TNF - α、IL - 1β、IL - 6和IL - 10的产生。如图3E-H所示,与对照组相比,LPS显著刺激了TNF - α、IL - 1β、IL - 6和IL - 10 mRNA的转录。SalC显著降低TNF - α、IL - 1β、IL - 6和IL - 10 mRNA的转录。 SalC/丹酚酸C抑制lps诱导的NF - κB活化[2] 免疫荧光结果显示,LPS刺激后p‑NF‑κB p65的核易位明显增加,而SalC可以抑制p‑NF‑κB p65的核易位(图4A)。Western blot法检测炎症相关蛋白的表达。活化形式的p‑NF‑κB p65在LPS刺激下显著增加(p < 0.01),SalC(0.1、1和5 μM)以浓度依赖的方式抑制LPS诱导的NF‑κB活化(p < 0.01,图4B、C)。图4 B、D表明p - IκBα水平在LPS组显著增加(p < 0.01),而SalC(0.1, 1和5 μM)下调p - IκBα水平显著(p < 0.05,p < 0.01和p < 0.01)。 SalC/丹酚酸C增加了BV2小胶质细胞中Nrf2和HO‑1的表达[2] Nrf2信号通路在炎症反应和氧化应激的调控中发挥重要作用。图5显示,LPS刺激组的细胞核Nrf2水平与正常对照组相比无显著差异(P > 0.05),Nrf2下游蛋白HO‑1的表达在两组之间也无显著差异(P > 0.05)。但SalC(0.1, 1和5 μM) Nrf2的表达明显增加(P < 0.01),以及其下游蛋白质HO - 1 (P < 0.05,P < 0.01和P < 0.01)。与LPS组相比,SalC(1和5 μM)也显著提高了抗氧化酶NQO1的表达(P < 0.01)。 SalC/丹酚酸C通过Nrf2通路抑制NF - κB活化[2] 阐明SalC是否通过Nrf2途径抑制NF - κB活化。使用siRNA技术敲除Nrf2。结果显示,Nrf2敲低逆转了SalC对TNF - α、IL - 1β、IL - 6和IL - 10产生的抑制作用(图6B-E)。此外,免疫荧光结果还显示,Nrf2敲低逆转了SalC对NF - κB核易位的抑制作用(图6A)。 AMPK/Nrf2信号通路参与了SalC/丹酚酸C对NF - κB 的抑制作用[2] 在信号通路研究中,我们使用特异的Nrf2 siRNA来阐明Nrf2与NF - κB p65之间的相互作用。利用AMPK抑制剂化合物C研究Nrf2上游蛋白。结果表明,复方C在BV2小胶质细胞和原代小胶质细胞中均逆转了SalC对Nrf2的激活作用。此外,化合物C预孵育也逆转了SalC对NF - κB的抑制作用。用Nrf2 siRNA处理可逆转SalC对NF - κB的抑制作用(图10)。上述结果表明,AMPK/Nrf2信号通路参与了SalC对NF - κB和神经炎症的抑制作用。 |
| 体内研究 (In Vivo) |
丹酚酸 C (20 mg/kg) 处理显着降低了逃避潜伏期。此外,与LPS模型组相比,丹酚酸C(10和20mg/kg)治疗组的平台穿越次数显着更高。与模型组相比,全身注射丹酚酸 C 可以降低大鼠脑组织 TNF-α、IL-1β 和 IL-6 水平。大脑皮层和海马体用于 iNOS 和 COX-2 水平的建模。丹参酚酸 C(5、10 和 20 mg/kg)治疗显着降低了皮质和海马的水平,而 p-AMPK、Nrf2、HO-1 和 NQO1 的水平呈剂量依赖性增加[2]。
SalC/丹酚酸C抑制大鼠神经炎症,改善记忆行为[2] 脂多糖诱导SD大鼠神经炎症。如图7A所示,Morris水迷宫实验中,LPS模型组大鼠的逃避潜伏期明显高于对照组(P < 0.01),而SalC(20 mg/kg)处理可显著降低大鼠的逃避潜伏期(P < 0.05)。此外,LPS模型组大鼠与对照组大鼠相比,穿越部位次数较少。与LPS模型组相比,SalC(10和20 mg/kg)处理显著增加了平台穿越次数(P < 0.05和P < 0.01,图7B)。上述结果表明,LPS模型组大鼠空间学习记忆功能受损,而SalC处理可减轻该损伤。与模型组相比,全身给药LPS可显著上调大鼠脑TNF - α、IL - 1β和IL - 6水平,而SalC可下调大鼠脑TNF - α、IL - 1β和IL - 6水平(图7C)。大鼠大脑皮层和海马区的iNOS和COX‑2水平高于对照组,而SalC处理显著下调了大鼠大脑皮层和海马区的iNOS和COX‑2水平(图7D-F)。进一步的免疫荧光研究表明,LPS增加了皮层和海马中GFAP和Iba1阳性细胞的荧光强度,SalC处理抑制了LPS诱导的神经炎症,降低了GFAP和Iba1的荧光强度(图7G-I)。 SalC/丹酚酸C激活AMPK/Nrf2,抑制NF - κB信号通路[2] 如图8所示,SalC(5、10和20 mg/kg)剂量依赖性地增加了大鼠大脑皮层和海马中p - AMPK、Nrf2、HO - 1和NQO1的水平。此外,全身给药LPS诱导NF - κB信号通路的激活,而SalC(5、10和20 mg/kg)剂量依赖性地抑制NF - κB p65磷酸化和i - κBα磷酸化(图9)。 |
| 酶活实验 |
酶动力学分析[1]
分别测定紫杉醇6-羟基化和阿斯咪唑o -去甲基阿斯咪唑对CYP2C8和CYP2J2的探针活性。制备了总体积为200 μL的培养混合物,培养混合物中有机溶剂的最终浓度小于1% (v/v)。孵育混合物如下:重组P450酶、nadph生成系统(10 mM葡萄糖6-磷酸、0.5 mM NADP、10 mM MgCl2、1单位葡萄糖6-磷酸脱氢酶)、100 mM磷酸钾缓冲液(pH 7.4)和一定浓度的探针底物。紫杉醇和阿斯咪唑在甲醇中溶解。在37 °C下进行5分钟的预孵育,然后加入nadph生成系统开始反应。根据初步实验(补充表1),所有孵育均在时间和蛋白浓度线性条件下进行。在培养混合物中加入不同浓度的紫杉醇(0.2 ~ 50 μmol· L−1)和阿斯咪唑(0.05 ~ 50 μmol· L−1)。使用Michaelis-Menten模型(GraphPad Prism 5, CA, USA)对酶活性-底物浓度数据进行非线性回归分析,确定了不同浓度的标记底物的Km(作用速率为其最大速度的一半时的底物浓度)和Vmax(给定条件下酶催化反应的最大初始速度)值。采用液相色谱-串联质谱(LC-MS/MS)测定培养混合物中代谢物的最终浓度。 筛选丹参类化合物对CYP2C8和CYP2J2的抑制作用[1] 孵育混合物中含有CYP2C8或CYP2J2、nadph生成系统、底物(终浓度 ~ Km)和单一测试化合物的不同浓度(0-50 μmol·L−1),在100 mM磷酸盐缓冲液(pH 7.4)中孵育。亲水组分(丹酚酸,SAs,例如丹酚酸C)溶解在水中,亲脂组分(TSTs)溶解在二甲亚砜中,其最终浓度小于混合物的1% (v/v)。以槲皮素和那那唑分别作为CYP2C8和CYP2J2的阳性对照。所有孵育均为三次,取平均值进行分析。 在抑制剂存在的情况下,CYP2C8和CYP2J2的活性以相应对照的百分比表示。从抑制百分比与逻辑抑制剂浓度的图中,通过使用非线性回归分析将剩余酶活性百分比(Y)与初始抑制剂浓度(X)数据的对数拟合到eq.(1)中,计算相应的IC50值(酶抑制50%所需的抑制剂浓度)。 (1)Y = 100/(1 + 10X−LogIC50) 抑制型研究[1] 当IC50值低于15 μmol·L−1时,测定丹参组分对特定CYPs的抑制类型和Ki值(抑制剂的抑制常数,等于酶/抑制剂复合物的解离常数)。采用紫杉醇(2.5、5和10 μmol·L−1)、阿斯米唑(0.5、1和2 μmol·L−1)和不同浓度的丹参成分构建Lineweaver-Burk和Dixon图,并估计其抑制类型和Ki值。其他程序与可逆抑制研究相似。 时间依赖性抑制研究[1] 当IC50值小于5 μmol·L−1时,采用传统的IC50移位法测定丹参组分对CYP2C8和CYP2J2的抑制作用。几种不同浓度的测试化合物与重组CYPs(2至10倍浓度)预孵育,有或没有nadph生成系统,在37 °C下,15 min。失活孵育后,将等分液(20-100 μL)转移到含有nadph生成系统和CYPs底物的新鲜孵育管(终体积200 μL)中。孵育、淬火、样品提取和分析程序与抑制型研究相同。 根据nadph生成系统的存在和不存在(式(5))确定的IC50值来计算IC50移位,明显的左移位(IC50移位bbbb2)表明存在基于机制的失活剂。 (5)IC50shift = IC50(-NADPH) / IC50(+NADPH) DI对CYP2C8 抑制作用的时间和浓度依赖性测定[1] 预孵育混合物中含有100 mmol·L−1磷酸钾缓冲液(pH 7.4)、重组CYP2C8(10 nmol·L−1)和各种浓度(0.5 ~ 8 μmol·L−1)的DI,总体积为100 μL。单独加入溶剂制备不含试验化合物的对照样品。在37 °C的预孵育液中,加入20 μL的nadph生成系统,继续孵育30 min。在孵育0、5、10、20和30 min时,收集50 μL的混合物,加入到由100 mM磷酸钾缓冲液(pH 7.4)、10 μmol·L−1紫杉醇和nadph生成系统组成的测定混合物(50 μL)中,用于测定紫杉醇羟化酶活性。将CYP2C8活性测定液在37 ℃下孵育20 min 后,加入100 μL乙腈制咪达唑仑终止反应。采用LC-MS/MS法测定6-羟基紫杉醇的浓度。 标记代谢物的测定[1] 采用部分验证的LC-MS/MS方法对紫杉醇的6-羟基化代谢物进行定量分析。重组CYP2C8体系在37 °C孵育后,加入100 μL的乙腈(内标)配制的20.16 ng·mL−1咪达唑仑停止反应。涡流混合1 min,然后以12,000 rpm离心10 min。将得到的上清液5 μL注入到由安捷伦1100系列高效液相色谱系统与API 4000质谱联用组成的LC-MS/MS系统中,该系统配备电喷雾电离(ESI)源进行离子生成。色谱柱为C18(1.8 μm, 100 × 4.6 mm;Agilent, USA)和分析物用乙腈(A)和0.1%甲酸水溶液(B)洗脱,流速为0.4 mL·min - 1,使用以下梯度程序:0-0.3 min, 60% A;0.3-2 min, 60%-90% A;2-4 min, 90% A;4-5.1min, 90%-60% a,柱温保持在40 °C。离子喷雾电压设置为5.0 kV进行正离子电离,加热器气体温度为550 ℃。氮气用作雾化气体(50psi)、辅助气体(55psi)和幕帘气体(15psi)。对6-羟基紫taxol m/z 870.4 ~ 286.3和咪达唑安定m/z 326.3 ~ 291.1的前体离子到产物离子的跃迁进行了多重反应监测(MRM)实验,前者的聚类势(DP)为60 V,碰撞能(CE)为−23 eV,后者的DP为60 V, CE为40 eV,没有内部干扰。6-羟基紫杉醇在1.17 ~ 117 nmolmL−1的浓度范围内呈线性。日内、日间精密度(CV%)分别小于2.7 ~ 8.2%和4.8 ~ 11.4%,准确度分别为96.2 ~ 109.5%和101.6 ~ 106.5%。在浓度为2.00、10.0和80.0 nmol·mL−1时,平均回收率为97.6 ~ 100.3%,基质效应为87.3 ~ 92.4%。在自动进样器条件下,6-羟基紫杉醇在处理后的样品中稳定(92.1-103.3%)至少24 h。 |
| 细胞实验 |
BV2小胶质细胞与原代大鼠小胶质细胞的培养及分组[2]
采用终浓度为1 μg/mL的LPS刺激建立细胞损伤模型。实验分为以下组:正常对照组(培养基),LPS刺激模型组(1 μg / mL有限合伙人),Salvianolic Acid C/ SalC低剂量组(1 μg / mL有限合伙人 + 0.1μM SalC), SalC中间剂量组(1 μg / mL有限合伙人 + 1 μM SalC),和SalC高剂量组(1 μg / mL有限合伙人 + 5 μM SalC)。BV2和原代小胶质细胞均用20 μM AMPK抑制剂化合物C预孵育1 h,然后用SalC处理。 MTT试验[2] MTT法测定Salvianolic Acid C /SalC对细胞活力的影响。首先,以1 × 104个细胞/孔的密度在96孔板上接种BV2小胶质细胞。然后将细胞与不同浓度的SalC和1 μg/mL LPS共孵育24 h。随后,移除上清液,并在每口井中加入MTT。孵育4 h后,每孔加入100 μL DMSO,在SpectraMax M5酶标仪上测定490 nm波长处的吸光度。 NO检测[2] 采用Griess法检测上清液中NO水平。实验结束时,将50 μL上清液转入新孔。然后加入50 μL Griess Reagent I混合。加入Griess试剂II 50 mL混合。最后,在540 nm处测量吸光度,计算NO浓度。 酶联免疫吸附试验(ELISA) [2] LPS刺激和Salvianolic Acid C /SalC孵育后,收集培养基上清液。炎症介质TNF - α、IL - 1β、IL - 6、IL - 10和PGE2的浓度根据生产厂家说明书采用相应的ELISA试剂盒检测。同时收集对照组、LPS模型组和LPS + SalC处理组大鼠脑组织中炎症介质TNF - α、IL - 1β、IL - 6浓度,并按上述说明书进行测定。 免疫荧光测定[2] 免疫荧光法观察iNOS和p‑NF‑κB p65的表达或细胞内定位。首先,将BV2小胶质细胞以1 × 每孔104个细胞的密度接种于96孔板上,孵育24 h。不同浓度Salvianolic Acid C/SalC与LPS共孵育24 h后,取出上清液。用4%多聚甲醛固定BV2小胶质细胞15 min。细胞用0.3% Triton X-100孵育10 min,随后用5% BSA阻断1 h,用抗iNOS和p‑NF‑κB p65一抗在4 °C孵育过夜。然后加入荧光二抗,孵育1.5 h。加入Hoechst 33258孵育30 min。最后,在Cellomics ArrayScan®VTI成像平台下获取荧光图像并进行测量 Nrf2 siRNA转染[2] 细胞接种于6孔板。24 h后,每孔加入0.8 mL siRNA转染培养基。与Nrf2 siRNA或对照siRNA孵育6 h后,再次加入正常培养基。细胞与不同浓度Salvianolic Acid C/SalC和1 μg/mL LPS孵育24 h。最后进行酶联免疫吸附试验、免疫荧光试验和免疫印迹法。 |
| 动物实验 |
系统性神经炎症动物模型及药物治疗[2]
Sprague-Dawley (SD) 大鼠(210-240 g)饲养于温度 21-24 °C、12 小时光照/12 小时黑暗循环、相对湿度 50% 的环境中。大鼠随机分为五组(每组 n = 12):对照组、LPS 模型组和 LPS + 丹酚酸 C (SalC) 治疗组(SalC 剂量分别为 5、10 和 20 mg/kg/天)。采用全身性 LPS 给药诱导大鼠神经炎症,该方法已被多项既往研究证实。大鼠系统性神经炎症的诱导方法参照既往报道。简而言之,为诱导系统性炎症,LPS 模型组大鼠腹腔注射溶于生理盐水的 LPS,剂量为 500 μg/kg,连续 7 天。在为期7天的实验期间,对照组大鼠仅腹腔注射生理盐水。LPS+SalC处理组大鼠在LPS注射后1小时,每天腹腔注射5、10或20 mg/kg剂量的SalC,持续7天。SalC注射后3小时进行Morris水迷宫实验。 |
| 参考文献 | |
| 其他信息 |
丹酚酸C属于苯并呋喃类化合物。
据报道,丹酚酸C存在于丹参、牛至以及其他有相关数据的生物体中。 中药和天然产物作为一种替代疗法或保健品,在中国和西方社会的使用日益普及。丹参(唇形科植物丹参的干燥根)富含酚酸和丹参酮,是一种广泛用于治疗心脑血管疾病的中药。本研究旨在探讨丹参中15种成分对CYP2C8和CYP2J2的抑制作用,这两种酶均在人肝脏和心血管系统中表达。本研究采用重组CYP2C8和CYP2J2,并确定了其抑制机制、动力学和类型。分别以紫杉醇6-羟基化和阿司咪唑O-去甲基阿司咪唑为探针,测定了CYP2C8和CYP2J2的活性。采用液相色谱-串联质谱法(LC-MS/MS)分析了代谢产物的生成。结果表明,丹酚酸A是CYP2C8的竞争性抑制剂(Ki = 2.5 μM),也是CYP2J2的混合型抑制剂(Ki = 7.44 μM)。丹酚酸C对CYP2C8(Ki = 4.82 μM)和CYP2J2(Ki = 5.75 μM)分别表现出中等程度的非竞争性和混合型抑制作用。丹参酮IIA是CYP2C8的中等程度的竞争性抑制剂(Ki = 1.18 μM)。二氢丹参酮I对CYP2J2具有中等程度的非竞争性抑制作用(Ki = 6.59 μM),但对CYP2C8具有机制性抑制作用(Ki = 0.43 μM,kinact = 0.097 min⁻¹)。丹参酮I是CYP2C8的中等程度的竞争性抑制剂(Ki = 4.20 μM)。这些结果表明,口服丹参制剂后,其通过CYP2C8介导的药物相互作用风险似乎不大;但当与其他药物联合口服时,其对肠道CYP2J2介导的药物代谢的抑制作用不容忽视。此外,本研究从CYP2C8和CYP2J2抑制的角度,为丹参成分的潜在药理机制提供了新的见解。 [1] 总之,我们的数据表明,丹参苷三碘甲状腺原氨酸(TI)、丹参苷二碘甲状腺原氨酸(TIIA)、丹参苷(Cry)、丹参苷二碘甲状腺原氨酸(DI)、丹参苷酸(SAA)、丹参苷酸(SAB)和丹参酚酸C/丹参苷酸(Salvianolic Acid C/SAC)对CYP2C8或CYP2J2具有不同程度的抑制作用,其IC50值均低于50 μmol·L−1。然而,本研究中测定的IC50/Ki值比健康人口服丹参制剂后这些化合物的血浆浓度高出50倍以上。因此,口服丹参制剂及其成分似乎不太可能引起CYP2C8介导的药物相互作用的显著风险;但是,当与其他药物联合使用时,不应忽视它们对肠道CYP2J2介导的药物代谢的抑制作用。此外,本研究从CYP2C8和CYP2J2抑制的角度,为丹参成分的潜在药理机制提供了新的见解,这对于深入理解丹参与药物代谢酶之间的相互作用非常有帮助。[1]神经退行性疾病与神经炎症密切相关。靶向炎症的药物已被证明在多种动物模型中有效。丹酚酸C (SalC) 是从丹参(Salvia miltiorrhiza Bunge)中分离得到的化合物,丹参是一种具有抑制炎症作用的植物。然而,SalC对LPS刺激的神经炎症的抗炎作用及其生物学机制尚不清楚。本文旨在研究其保护作用及其抗炎机制。本研究采用LPS在体内和体外诱导SD大鼠和小胶质细胞的神经炎症,并采用MTT法检测细胞活力。采用ELISA法检测TNF-α、IL-1β、IL-6、IL-10和PGE2的水平。采用Western blot分析检测p-AMPK、p-NF-κB p65、p-IκBα、Nrf2、HO-1和NQO1蛋白的表达。采用免疫荧光法研究NF-κB p65的核转位。使用特异性Nrf2 siRNA阐明Nrf2与NF-κB p65的相互作用。使用AMPK抑制剂Compound C研究Nrf2的上游蛋白。结果表明,LPS诱导大鼠脑组织和小胶质细胞中炎症细胞因子的过表达,并介导NF-κB p65的磷酸化和核转位。SalC逆转LPS诱导的炎症反应,并抑制NF-κB的激活。 SalC 还上调了 p-AMPK、Nrf2、HO-1 和 NQO1 的表达。但 SalC 的抗炎和 NF-κB 抑制作用可通过转染特异性 Nrf2 siRNA 或使用强效 AMPK 抑制剂 Compound C 进行干扰而减弱。总之,SalC 通过激活 AMPK/Nrf2 信号通路,在体内和体外均抑制了 LPS 诱导的炎症反应和 NF-κB 活化。[2] 综上所述,上述结果表明,丹酚酸 C/SalC 通过抑制 NF-κB 信号通路来抑制 LPS 诱导的炎症细胞因子的产生。而 NF-κB 的抑制是由 AMPK/Nrf2 信号通路触发的。[2] |
| 分子式 |
C26H20O10
|
|---|---|
| 分子量 |
492.4310
|
| 精确质量 |
492.105
|
| CAS号 |
115841-09-3
|
| PubChem CID |
13991590
|
| 外观&性状 |
Light yellow to yellow solid powder
|
| 密度 |
1.6±0.1 g/cm3
|
| 沸点 |
844.2±65.0 °C at 760 mmHg
|
| 闪点 |
464.4±34.3 °C
|
| 蒸汽压 |
0.0±3.3 mmHg at 25°C
|
| 折射率 |
1.752
|
| LogP |
3.12
|
| tPSA |
177.89
|
| 氢键供体(HBD)数目 |
6
|
| 氢键受体(HBA)数目 |
10
|
| 可旋转键数目(RBC) |
8
|
| 重原子数目 |
36
|
| 分子复杂度/Complexity |
802
|
| 定义原子立体中心数目 |
1
|
| SMILES |
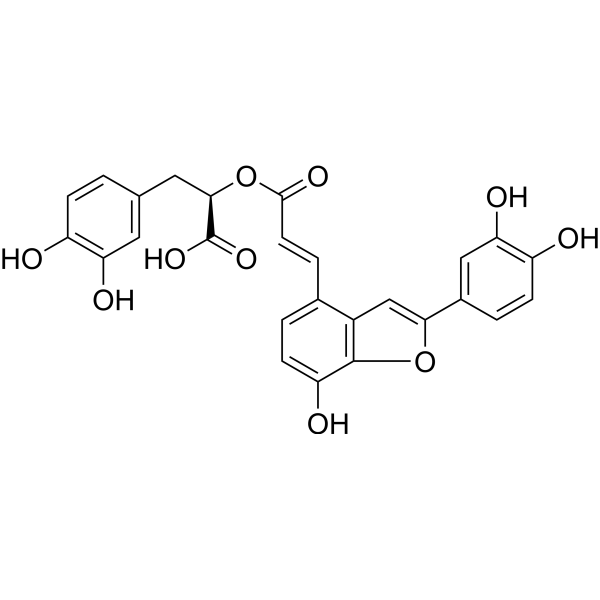
C1=CC(=C(C=C1C[C@H](C(=O)O)OC(=O)/C=C/C2=C3C=C(OC3=C(C=C2)O)C4=CC(=C(C=C4)O)O)O)O
|
| InChi Key |
GCJWPRRNLSHTRY-VURDRKPISA-N
|
| InChi Code |
InChI=1S/C26H20O10/c27-17-5-1-13(9-20(17)30)10-23(26(33)34)35-24(32)8-4-14-2-7-19(29)25-16(14)12-22(36-25)15-3-6-18(28)21(31)11-15/h1-9,11-12,23,27-31H,10H2,(H,33,34)/b8-4+/t23-/m1/s1
|
| 化学名 |
(2R)-3-(3,4-dihydroxyphenyl)-2-[(E)-3-[2-(3,4-dihydroxyphenyl)-7-hydroxy-1-benzofuran-4-yl]prop-2-enoyl]oxypropanoic acid
|
| 别名 |
Salvianolic acid C; 115841-09-3; UNII-I16H9Z53ZL; I16H9Z53ZL; (2R)-3-(3,4-dihydroxyphenyl)-2-[(E)-3-[2-(3,4-dihydroxyphenyl)-7-hydroxy-1-benzofuran-4-yl]prop-2-enoyl]oxypropanoic acid; Benzenepropanoic acid, alpha-(((2E)-3-(2-(3,4-dihydroxyphenyl)-7-hydroxy-4-benzofuranyl)-1-oxo-2-propen-1-yl)oxy)-3,4-dihydroxy-, (alphaR)-; Benzenepropanoic acid, alpha-[[(2E)-3-[2-(3,4-dihydroxyphenyl)-7-hydroxy-4-benzofuranyl]-1-oxo-2-propen-1-yl]oxy]-3,4-dihydroxy-, (alphaR)-; BENZENEPROPANOIC ACID, .ALPHA.-(((2E)-3-(2-(3,4-DIHYDROXYPHENYL)-7-HYDROXY-4-BENZOFURANYL)-1-OXO-2-PROPEN-1-YL)OXY)-3,4-DIHYDROXY-, (.ALPHA.R)-;
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month 注意: 请将本产品存放在密封且受保护的环境中(例如氮气保护),避免吸湿/受潮和光照。 |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO : ~50 mg/mL (~101.54 mM)
|
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.5 mg/mL (5.08 mM) (饱和度未知) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 25.0 mg/mL澄清DMSO储备液加入到400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: ≥ 2.5 mg/mL (5.08 mM) (饱和度未知) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 例如,若需制备1 mL的工作液,可将 100 μL 25.0 mg/mL澄清DMSO储备液加入900 μL 20% SBE-β-CD生理盐水溶液中,混匀。 *20% SBE-β-CD 生理盐水溶液的制备(4°C,1 周):将 2 g SBE-β-CD 溶解于 10 mL 生理盐水中,得到澄清溶液。 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.0307 mL | 10.1537 mL | 20.3075 mL | |
| 5 mM | 0.4061 mL | 2.0307 mL | 4.0615 mL | |
| 10 mM | 0.2031 mL | 1.0154 mL | 2.0307 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 463611831
463611831
