| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 1mg |
|
||
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| Other Sizes |
|

| 靶点 |
Natural product; anticancer; anthelmintic; Notch
|
|---|---|
| 体外研究 (In Vitro) |
治疗 48 小时后,马钱子碱 D 对 H460 和 A549 细胞活力产生剂量依赖性影响,IC50 值分别为 0.5 和 0.6 μM [1]。在 H460 和 A549 细胞中,马钱子碱 D (0.125-1.0 μM) 剂量调节可增强染色质凝聚、膜联蛋白 V 诱导的细胞聚集和 caspase 细胞死亡 [1]。
马钱子碱D(BD)显示出对DBM的强效抗饲料活性[1] 鉴定了Bruceine D (BD),并通过根和叶施用评估了其保护开花大白菜免受DBM幼虫取食的能力。据报道,印楝素是一种具有系统作用的卓越的喂养威慑剂;因此,采用印楝素进行比较。 通过根施用进行无选择饲喂试验[1] 在24和48小时与100μg/mL的马钱子碱D/Bruceine D (BD)或印楝素孵育后,评估了对DBM三龄幼虫的拒食活性(图2A)。在根上施用马钱子碱D后,大白菜叶片表现出优异的拒食活性,分别减少了93.80%和96.83%的消耗。与马钱子碱D相比,印楝素通过这种应用是无效的,在治疗24小时后没有显示出进食抑制作用,在治疗48小时后的拒食率为22.60%。然后,通过HPLC-HRESIMS定量叶片中马钱子碱D和印楝素的含量(图2C和D)。马钱子碱D和印楝素都具有显著的系统行为,并且随着时间的推移不断转移到叶片中(图2B)。根系施用24和48小时后,马钱子碱D的浓度分别为38.69μg/g(鲜重,FW)和108.45μg/g(FW),显著高于印楝素21.43μg/g(FWs)和36.88μg/g(FWD)的浓度(p<0.05)。显然,在48小时内,马钱子碱D比印楝素表现出更好的全身特性。叶片中马钱子素D的剂量足以防止DBM的摄食行为,而印楝素对摄入量的影响很小。 通过叶面施用进行无选择饲喂试验[1] Bruceine D(BD)显示出强烈的拒食作用。为了评估对DBM三龄幼虫的拒食活性,使用2.5至160μg/mL的一系列浓度覆盖开花的大白菜叶盘,并使用印楝素作为阳性对照,不同浓度为20、40、80、160、320、640μg/mL。饲喂24小时后,这两种化合物表现出剂量依赖的拒食活动行为。马钱子碱D和印楝素的AFC50值分别为0.11和0.68μg/cm2,这表明在我们的实验条件下,马钱子素D的拒食活性比印楝素强约6.2倍。 Bruceine D(BD)在体外抑制HCC细胞活力并诱导细胞凋亡[2] 为了鉴定HCC的治疗药物,我们试图在HEK293T细胞中发现一种Notch抑制剂,该细胞含有基于萤光素酶的RBP-Jκ-Luc报告子,该报告子具有ICN(Notch的细胞内结构域)驱动的萤光素酶,以响应Notch信号活性(图S1A)。在1600种天然产物的筛选中,BD对RBP-Jκ-Luc活性的抑制作用最强(图S1B)。为了确定BD是否是HCC抑制剂,将Huh7和Hep3B细胞用不同浓度的BD处理48小时。CCK8检测显示,这两种细胞类型的增殖都受到强烈的剂量和时间依赖性抑制,并且HCC细胞的敏感性高于人肝细胞L-02(图S2A,B)。与对细胞存活的影响类似,BD还以浓度依赖性方式抑制Huh7和Hep3B的癌症细胞集落形成(图S2C)。此外,与对照细胞相比,BD诱导Huh7和Hep3B细胞凋亡的剂量依赖性增加,尤其是早期凋亡(图S2D)。这些结果证实,BD对Notch信号的抑制是通过抑制HCC细胞的增殖和诱导凋亡来实现的。 Bruceine D (BD)抑制HCC细胞中Jagged1的蛋白质合成[2] 为了了解BD抑制Notch信号传导的潜在机制,我们接下来确定了BD对Huh7和Hep3B细胞系中Notch信号相关蛋白的影响。值得注意的是,虽然我们观察到Notch 1、2或3没有变化,但Notch配体Jagged1、切割的Notch受体NICD和Notch特异性下游蛋白Hes1都明显受到BD的下调(图4A)。 Jagged1是β-catenin靶基因,是Bruceine D (BD)介导的HCC增殖抑制所必需的 Jagged1的抑制与恶性HCC中肿瘤负荷的减少有关。为了说明Jagged1与BD作用的关系,我们在HCC细胞中进行了Jagged1的功能获得和丧失测定(图5A)。台盼蓝排斥试验表明,尽管BD具有细胞毒性作用,但含有丰富Jagged1的Huh7细胞仍显示出增殖能力(图5B)。与BD抑制的效果一致,与对照组相比,Jagged1的敲除导致细胞存活率降低和凋亡增加(图5B)。此外,接受BD联合shJagged1治疗的组与单独接受shJagged2治疗的组没有差异。这些结果表明,BD的作用需要Jagged1。 Bruceine D(BD)诱导β-catenin的蛋白酶体依赖性降解[2] 我们进一步推测BD是否通过Wnt通路下调Jagged1的表达。为了研究这一假设,我们测试了BD对HCC细胞中β-catenin/Tcf4转录复合物的影响。Huh7细胞与含有3个串联Tcf共有结合位点(TOP)或突变Tcf结合位点(FOP)的萤光素酶报告基因和作为正常转染对照的Renilla萤光素酶报道基因共转染。根据我们的假设,BD以剂量依赖的方式有效地抑制了TOP萤光素酶的活性,但没有抑制FOP萤光素酶(图6A)。 |
| 体内研究 (In Vivo) |
在鸦胆子中鉴定出一种类Bruceine D (BD)。对小菜蛾(DBM,Plutella xylostella L.)、甜菜夜蛾(Spodoptera exigua Hübner)和棉花叶虫(Spodopera litura Fabricius)表现出优异的系统性和拒食活性。其对DBM三龄幼虫的拒食作用比印楝素强约6.2倍。当在根部以100μg/mL的浓度施用Bruceine D 24和48小时时,其在开花大白菜(Brassica campestris L.ssp.chinensis var.utiliz Tsen et Lee)叶片中的浓度分别为38.69μg/g(鲜重,FW)和108.45μg/g(FW)。这些浓度可以达到93.80%和96.83%的拒食效果,明显高于印楝素。与印楝素相似,Bruceine D对昆虫幼虫也有强烈的生长抑制作用。在喂食20μg/g的Bruceine D后,没有观察到蛹。结果表明,Bruceine D是一种有效的植物性昆虫拒食剂,具有出色的系统性,具有强大的害虫生长抑制活性[1]。
本研究旨在确定鸦胆子干果的驱虫活性,并分离和表征其活性成分。从B.javanica果实中提取的甲醇提取物对Dactylogyrus intermedius显示出显著的驱虫活性(EC(50)(中效浓度)值=49.96 mg l(-1))。基于这一发现,甲醇提取物在硅胶柱色谱法上进行生物测定指导的分级,得到两种已知的具有强效活性的奎西诺,即Bruceine a和Bruceine D (BD)。Bruceinea和D对中间型D.表现出显著的活性,EC(50)值分别为0.49 mg l(-1)和0.57 mg l(-1),比阳性对照甲苯咪唑更有效(EC(50”值=1.25 mg l(-1-))。此外,BruceineA和D对宿主(鲫鱼)的48小时中位致死浓度(LC(50))比中间D.的EC(50)高10.6倍和9.7倍。这些结果提供了证据,表明分离的化合物可能是控制Dactylogyrus的新型抗寄生虫药物的潜在来源。这是首次报道爪哇小蠊对间日疟原虫的体内驱虫研究。 |
| 酶活实验 |
肝细胞癌(HCC)以高死亡率和有限的治疗方法而闻名。Wnt和Notch信号通路的异常激活对肝癌的发生和进展至关重要。在这里,我们使用RBP-Jκ依赖的萤光素酶报告系统鉴定了一种小分子,即马钱子碱D(BD),作为Notch抑制剂。BD显著抑制了肝肿瘤的生长,并增强了索拉非尼在各种小鼠HCC模型中的治疗效果。从机制上讲,BD促进β-catenin的蛋白酶体降解及其核积累的耗竭,这反过来又破坏了HCC中Notch配体Jagged1的Wnt/β-catenin-dependent转录。我们的研究结果提供了关于一种新型Wnt/Notch串扰抑制剂的重要信息,该抑制剂与索拉非尼协同治疗HCC,因此具有很高的临床影响[2]。
|
| 细胞实验 |
细胞活力测定[2]
将细胞在96孔板中培养过夜(3000个细胞/孔),然后用指定浓度的Bruceine D(BD)或索拉非尼处理24、48或72小时。然后,在每个孔上用10μl细胞计数试剂盒-8(CCK8)处理细胞1小时。在450nm处测量光密度,并将其归一化为无细胞培养基的背景吸光度。所有样品均进行了三次化验。 软琼脂集落形成试验和克隆形成试验[2] 将6孔板中的细胞(1000/孔)在含有10%FBS的1 ml 0.33%基础培养基鹰琼脂或含有10%FBS、浓度为3.5 ml 0.5%基础培养基的3.5 ml鹰琼脂中与或不与Bruceine D(BD)(0-1μM)一起孵育。在5%CO2的培养箱中,将培养物在37°C下保持10-14天。使用Image Pro Plus软件程序在显微镜下计数细胞集落。 流式细胞术分析细胞凋亡 用Annexin V-FITC凋亡检测试剂盒检测细胞凋亡。在用膜联蛋白V-FITC和碘化丙啶(PI)染色之前,用马钱子碱D(BD)或索拉非尼单独或联合处理细胞24小时。在室温下在黑暗中孵育5分钟后,通过流式细胞术分析细胞凋亡。 台盼蓝排斥试验[2] Huh7细胞在6孔板中培养过夜,与或不与Bruceine D(BD)一起孵育24小时。根据推荐的方案用台盼蓝染料对细胞进行染色。 免疫细胞化学(ICC)和免疫组织化学(IHC)[2] 细胞被放置在35毫米的玻璃板上,并在用Bruceine D(BD)(0.5-1μM)处理24小时之前生长过夜。DMSO用作未处理的对照。将第一代β-catenin抗体稀释1:100,并在4°C下孵育过夜。然后,将1:200稀释的Alexa Fluor 594标记的山羊抗兔IgG二抗加入固定的Huh7细胞中。然后将带有细胞的盖玻片安装在含有DAPI染色(CST)的Vectashield安装介质的载玻片上。荧光成像由LAS AF Lite软件和Leica TCS SP8 X显微镜捕获 细胞转染[2] 使用X-tremeGENE HP DNA转染试剂在Opti-MEM培养基中用萤光素酶报告质粒和pcDNA4/TO-N100-GV瞬时转染T-REx-HeLa细胞。转染24小时后,在受试化合物存在下用1μg/ml四环素处理细胞。孵育24小时后,将细胞在Glo裂解缓冲液中裂解5分钟,并通过在96孔Nunc MaxiSorp平板中混合裂解物和Bright Glo萤光素酶测定试剂来测量萤光素酶活性。在室温下孵育5分钟后,在微孔板读数器中监测单个孔中的发光。 在Opti-MEM培养基中,用Notch1-NEXT质粒或pcDNA3质粒(对照)以及2μl X-tremeGENE HP DNA转染试剂转染HEK293细胞。转染24小时后,将HEK293细胞与DAPT(GSI-IX)或Bruceine D(BD)一起孵育24小时,提取全细胞进行蛋白质印迹分析。 根据制造商的建议,用shCTNNB1和shJAG1瞬时转染Huh7细胞,并用lipo2000转染。转染24小时后,用DMSO或Bruceine D(BD)处理细胞24小时,提取全细胞进行蛋白质印迹。 萤光素酶报告物测定[2] 简而言之,将细胞接种到96孔板中,并与pTOPflash或pFOPflash载体以及肾荧光素酶报告载体pRL-TK共转染,作为转染效率的对照。转染12小时后,将细胞与Bruceine D(BD)一起孵育24小时。根据推荐的方案测量萤光素酶和肾菌的活性并使其标准化。为了进行药物筛选,将含有RBP-JκReporter的HEK293细胞系接种到96孔板中。孵育过夜后,将1600种10μM的天然产物加入每个孔中24小时,使用Promega E1500试剂盒检测萤光素酶活性。 |
| 动物实验 |
昆虫生长调节试验[1]
将布鲁辛D溶解于二甲基亚砜(DMSO)中,浓度为5000 μg/mL,然后添加到小菜蛾幼虫饲料中,最终浓度分别为2.5、5、10和20 μg/g。将刚蜕皮的三龄幼虫(每组n = 100)放入昆虫饲养容器中。饥饿3小时后,提供充足的食物。以饲喂含DMSO饲料的幼虫作为对照组。分别在饲喂含10和20 μg/g布鲁辛D饲料后6、12、24和48小时,测量幼虫的体重和饲料消耗量。每隔12小时统计处理组和对照组的化蛹和羽化个体数量。同时,观察了幼虫的发育过程和表型。 体内驱虫药效测定[3] 测定了各组分和纯化合物(如布鲁辛D (BD))对中间杜氏线虫(D. intermedius)的驱虫活性。试验在5升容量的玻璃水箱中进行,水箱内装有2升曝气地下水,每个水箱中分别放入待测样品和5条先前感染的鱼。水的pH值范围为7.0至7.5,溶解氧浓度为6.2至7.8 mg l−1(饱和度为72%至85%),水温恒定在24 ± 1 °C。进行初步试验是为了确定适中的浓度范围,以避免高浓度导致鱼类死亡。将衍生组分和纯化合物溶解于1 mL二甲基亚砜(DMSO)中,并用蒸馏水定容至100 mL,用于配制不同浓度的测试溶液。甲醇提取物和分离组分分别根据初始测试结果,设置不同的浓度系列。除纯化合物1和2进行三次重复实验外,所有实验均独立重复两次。纯化合物1和2的实验方案与爪哇苦参(B. javanica)组分的实验方案相同,使用五个浓度:0.5、1.0、1.5、2.0和2.5 mg/L。为排除DMSO对寄生虫的潜在影响,设置阴性对照,使用含有相应百分比DMSO的蒸馏水作为测试样品;甲苯咪唑作为阳性对照,使用不同浓度系列:0.6、1.0、1.5、2.0和2.5 mg/L。 48小时后,所有处理组和对照组中存活的金鱼均被实施脊髓切断术进行安乐死,以便进行尸检。将鳃片置于载玻片上,在4×10倍光学显微镜下计数鳃上的寄生虫数量,以确定每条感染金鱼的平均寄生虫数量。 急性毒性试验[3] 使用健康金鱼测定纯化合物(1和2)/布鲁辛D(BD)的急性毒性。试验重复两次,每次使用10条金鱼,置于5升容量的水箱中,每个水箱装有2升试验溶液,试验温度为24±1℃。根据初始试验结果,纯化合物的浓度设置成不同的系列。对照组在相同的试验条件下进行,但不添加任何化学物质。当金鱼鳃盖不再活动且对机械刺激(玻璃棒触碰)无反应时,记录其死亡。所有观察到的死鱼均及时从培养基中移除,以避免水质恶化。暴露48小时后,记录处理组和对照组的鱼死亡率。 皮下HCC异种移植模型研究[2] 四周龄雄性Balb/c裸鼠饲养于特定病原体清除(SPF)条件下。体内研究中,将5 × 10⁶个Huh7细胞悬浮于100 μl PBS中,皮下注射至小鼠右侧腹部。根据预实验数据,将小鼠随机分为3组(每组n=6),分别通过尾静脉注射给予0.9%氯化钠生理盐水(赋形剂)或布鲁辛D(BD),剂量分别为0.75 mg/kg和1.5 mg/kg体重,每日一次,连续10天。肿瘤体积按以下公式计算:肿瘤体积(mm3)= 1/2 ×(肿瘤长度)×(肿瘤宽度)。 |
| 参考文献 |
[1]. Bruceine D Isolated from Brucea Javanica (L.) Merr. as a Systemic Feeding Deterrent for Three Major Lepidopteran Pests. J Agric Food Chem. 2019 Apr 17;67(15):4232-4239.
[2]. Bruceine D inhibits hepatocellular carcinoma growth by targeting β-catenin/jagged1 pathways. Cancer Lett. 2017 Sep 10;403:195-205. [3]. In vivo anthelmintic activity of bruceine A and bruceine D from Brucea javanica against Dactylogyrus intermedius (Monogenea) in goldfish (Carassius auratus). Vet Parasitol. 2011 Apr 19;177(1-2):127-33. |
| 其他信息 |
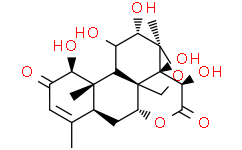
Bruceine D 是一种苦木素类化合物,其结构为 13,20-环氧苦木-3-烯,在 1、11、12、14 和 15 位被羟基取代,在 2 和 16 位被羰基取代。它从毛叶苦木(Brucea mollis)茎的乙醇提取物中分离得到,具有细胞毒活性。它可作为代谢物、抗肿瘤剂和植物代谢物发挥作用。它是一种δ-内酯、戊醇、苦木素类化合物、有机杂五环化合物和仲α-羟基酮。它来源于苦木烷的氢化物。
据报道,毛叶苦木(Brucea mollis)、印度苦木(Samadera indica)和爪哇苦木(Brucea javanica)中均含有 Bruceine D,并有相关数据。 内吸性是杀虫剂的理想特性。许多植物化学物质具有良好的内吸性,因此是新型内吸性杀虫剂成分的天然来源。从苦参(Brucea javanica (L.) Merr.)中鉴定出苦参素类化合物布鲁辛D,它对小菜蛾(Plutella xylostella L.)、甜菜夜蛾(Spodoptera exigua Hübner)和棉铃虫(Spodoptera litura Fabricius)表现出优异的内吸性和拒食活性。其对小菜蛾三龄幼虫的拒食效果约为印楝素的6.2倍。当以100 μg/mL的浓度将布鲁辛D施用于根部24小时和48小时后,开花期大白菜(Brassica campestris L. ssp. chinensis var. utiliz Tsen et Lee)叶片中布鲁辛D的浓度分别为38.69 μg/g(鲜重)和108.45 μg/g(鲜重)。这些浓度可分别达到 93.80% 和 96.83% 的拒食效果,显著高于印楝素。与印楝素类似,布鲁辛 D 也对昆虫幼虫具有显著的生长抑制作用。饲喂 20 μg/g 布鲁辛 D 后,未观察到蛹的形成。结果表明,布鲁辛 D 是一种有效的植物性昆虫拒食剂,具有优异的内吸性,可显著抑制害虫的生长。[1] 肝细胞癌 (HCC) 以高死亡率和有限的治疗方法而闻名。Wnt 和 Notch 信号通路的异常激活是肝癌发生和发展的关键。本研究利用 RBP-Jκ 依赖性荧光素酶报告系统,鉴定出一种小分子化合物布鲁辛 D (BD) 为 Notch 抑制剂。BD 显著抑制了肝肿瘤的生长,并在多种小鼠 HCC 模型中增强了索拉非尼的治疗效果。从机制上讲,BD促进β-catenin的蛋白酶体降解及其在细胞核内的积累,进而破坏肝细胞癌(HCC)中Notch配体Jagged1的Wnt/β-catenin依赖性转录。我们的研究结果为一种新型Wnt/Notch信号通路抑制剂提供了重要信息,该抑制剂与索拉非尼具有协同作用,可用于治疗HCC,因此具有重要的临床应用价值。[2] 本研究旨在确定爪哇番荔枝(Brucea javanica)干燥果实的驱虫活性,并分离和鉴定其活性成分。爪哇番荔枝果实的甲醇提取物对中间指环虫(Dactylogyrus intermedius)表现出显著的驱虫活性(EC50值(半数有效浓度)=49.96 mg l(-1))。基于此发现,采用生物活性导向的分级分离方法,通过硅胶柱色谱法对甲醇提取物进行分离,得到两种已知的具有强效活性的苦木素类化合物:布鲁辛A和布鲁辛D。布鲁辛A和D均对中间指环虫(D. intermedius)表现出显著的活性,其EC50值分别为0.49 mg/L和0.57 mg/L,优于阳性对照药物甲苯咪唑(EC50值为1.25 mg/L)。此外,布鲁辛A和D对宿主(金鱼,Carassius auratus)的48小时半数致死浓度(LC50)分别是其对中间指环虫EC50值的10.6倍和9.7倍。这些结果表明,分离得到的化合物可能是控制指环虫的新型抗寄生虫药物的潜在来源。这是关于对 B. javanica 进行体内驱虫研究以对抗 D. intermedius 的第一份报告。[3] |
| 分子式 |
C20H26O9
|
|---|---|
| 分子量 |
410.4150
|
| 精确质量 |
410.157
|
| 元素分析 |
C, 58.53; H, 6.39; O, 35.08
|
| CAS号 |
21499-66-1
|
| 相关CAS号 |
21499-66-1
|
| PubChem CID |
122784
|
| 外观&性状 |
White to off-white solid powder
|
| 密度 |
1.6±0.1 g/cm3
|
| 沸点 |
661.3±55.0 °C at 760 mmHg
|
| 闪点 |
235.8±25.0 °C
|
| 蒸汽压 |
0.0±4.5 mmHg at 25°C
|
| 折射率 |
1.670
|
| LogP |
-0.5
|
| tPSA |
153.75
|
| 氢键供体(HBD)数目 |
5
|
| 氢键受体(HBA)数目 |
9
|
| 可旋转键数目(RBC) |
0
|
| 重原子数目 |
29
|
| 分子复杂度/Complexity |
855
|
| 定义原子立体中心数目 |
11
|
| SMILES |
O1C([H])([H])[C@@]23C4([H])C([H])([H])C5([H])C(C([H])([H])[H])=C([H])C(C([H])([C@]5(C([H])([H])[H])C2([H])C([H])(C([H])([C@]1(C([H])([H])[H])[C@@]3(C([H])(C(=O)O4)O[H])O[H])O[H])O[H])O[H])=O
|
| InChi Key |
JBDMZGKDLMGOFR-KQSRGDCESA-N
|
| InChi Code |
InChI=1S/C20H26O9/c1-7-4-9(21)13(23)17(2)8(7)5-10-19-6-28-18(3,14(24)11(22)12(17)19)20(19,27)15(25)16(26)29-10/h4,8,10-15,22-25,27H,5-6H2,1-3H3/t8-,10+,11+,12+,13+,14-,15-,17-,18+,19+,20+/m0/s1
|
| 化学名 |
(1R,2R,3R,6R,8S,12S,13S,14R,15R,16S,17R)-2,3,12,15,16-pentahydroxy-9,13,17-trimethyl-5,18-dioxapentacyclo[12.5.0.01,6.02,17.08,13]nonadec-9-ene-4,11-dione
|
| 别名 |
Bruceine-D; Bruceine D; 21499-66-1; Brucein D; CHEBI:68931; C08752; (1R,2R,3R,6R,8S,12S,13S,14R,15R,16S,17R)-2,3,12,15,16-Pentahydroxy-9,13,17-trimethyl-5,18-dioxapentacyclo[12.5.0.01,6.02,17.08,13]nonadec-9-ene-4,11-dione; (1R,2S,3R,3aR,3a1R,4R,6aR,7aS,11S,11aS,11bR)-1,2,3a,4,11-pentahydroxy-3,8,11a-trimethyl-1,2,3,3a,4,7,7a,11,11a,11b-decahydro-5H-3,3a1-(epoxymethano)dibenzo[de,g]chromene-5,10(6aH)-dione; AC1L9BNP; Bruceine D
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month 注意: 本产品在运输和储存过程中需避光。 |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO: 82~100 mg/mL (199.8~243.7 mM)
H2O: ~5 mg/mL (~12.2 mM) |
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.5 mg/mL (6.09 mM) (饱和度未知) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 25.0 mg/mL澄清DMSO储备液加入到400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: ≥ 2.5 mg/mL (6.09 mM) (饱和度未知) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 例如,若需制备1 mL的工作液,可将 100 μL 25.0 mg/mL澄清DMSO储备液加入900 μL 20% SBE-β-CD生理盐水溶液中,混匀。 *20% SBE-β-CD 生理盐水溶液的制备(4°C,1 周):将 2 g SBE-β-CD 溶解于 10 mL 生理盐水中,得到澄清溶液。 View More
配方 3 中的溶解度: ≥ 2.5 mg/mL (6.09 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.4365 mL | 12.1826 mL | 24.3653 mL | |
| 5 mM | 0.4873 mL | 2.4365 mL | 4.8731 mL | |
| 10 mM | 0.2437 mL | 1.2183 mL | 2.4365 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
 BBO-11818
BBO-11818
 RMC-5127
RMC-5127
 Degarelix acetate hydrate
Degarelix acetate hydrate
 DDO-2728
DDO-2728
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 COA
COA
 463611831
463611831
