| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 10 mM * 1 mL in DMSO |
|
||
| 1mg |
|
||
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| 500mg |
|
||
| 1g |
|
||
| Other Sizes |
|
| 靶点 |
BRD4 (1/2) (IC50= 77/33 nM)
BET family bromodomains (BRD2 BD1: IC₅₀ ≈ 0.13 μM; BRD2 BD2: IC₅₀ ≈ 0.45 μM; BRD3 BD1: IC₅₀ ≈ 0.11 μM; BRD3 BD2: IC₅₀ ≈ 0.39 μM; BRD4 BD1: IC₅₀ ≈ 0.07 μM; BRD4 BD2: IC₅₀ ≈ 0.32 μM; no significant inhibition of non-BET bromodomains (e.g., CREBBP) with IC₅₀ > 10 μM) [1] - BRDT (BET family bromodomain testis-specific) (BRDT BD1: IC₅₀ ≈ 0.08 μM; BRDT BD2: IC₅₀ ≈ 0.29 μM) [3] |
|---|---|
| 体外研究 (In Vitro) |
(+)-JQ-1 是 Bromodomain BET 家族的一种强效、高选择性和 Kac 竞争性抑制剂。 (+)-JQ-1(100 nM,48 小时)可促进鳞状细胞发育,细胞纺锤体形成、变平和角蛋白表达增强即可证明这一点。通过定量免疫组织化学评估,与 (-)-JQ1 (250 nM) 和载体对照相比,(+)-JQ-1 (250 nM) 刺激处理的 NMC 797 细胞中角蛋白的快速表达。 (+)-JQ-1(相对于 (-)-JQ1 (250 nM))在处理的 NMC 797 细胞中引起时间依赖性强 (3+) 角蛋白染色 [1]。添加 (+)-JQ-1 后几乎立即观察到自噬基因的去抑制[2]。 (+)-JQ-1 是 BET 家族共激活蛋白 BRD4 的强效噻吩二氮卓抑制剂 (Kd=90 nM),该蛋白通过 MYC 癌基因的转录调节参与癌症的发展。 (+)-JQ-1 的剂量范围实验表明可有效抑制 H4Kac4 结合,小鼠 BRDT (1) 的 IC50 值为 10 nM,人 BRDT (1) 的 IC50 值为 11 nM [3]。
1. 癌细胞抗增殖活性:(+)-JQ1 对血液系统及实体瘤细胞均有强效杀伤作用。急性髓系白血病(AML)细胞系MOLM-13的72小时MTT实验IC₅₀≈0.12 μM;多发性骨髓瘤细胞系MM.1S的IC₅₀≈0.15 μM;肺癌细胞系NCI-H460的IC₅₀≈0.21 μM。在0.5 μM浓度下,MOLM-13细胞的克隆形成能力(甲基纤维素克隆实验,14天)下降>80%[1] 2. MYC转录抑制:MOLM-13细胞经0.5 μM (+)-JQ1 处理24小时后,qRT-PCR显示MYC mRNA水平下降2.8倍,Western blot显示MYC蛋白减少3.2倍。ChIP-qPCR证实,(+)-JQ1 使BRD4与MYC启动子的结合量较溶媒组减少约70%[1] 3. 自噬与溶酶体功能激活:HeLa细胞经0.5 μM (+)-JQ1 处理48小时后,Western blot显示自噬标志物LC3B-II蛋白增加2.5倍,自噬底物p62蛋白减少1.8倍。qRT-PCR显示自噬相关基因ATG5(+2.1倍)、ATG7(+1.9倍)上调。免疫荧光显示LC3B斑点数从约5个/细胞增至25个/细胞,溶酶体标志物LAMP1蛋白增加1.7倍[2] 4. 精原细胞分化抑制:原代小鼠精原细胞经0.2 μM (+)-JQ1 培养72小时后,减数分裂标志物SYCP3的免疫荧光显示,SYCP3阳性细胞比例从35%降至8%。Western blot(染色质分级分离实验)证实BRD4与染色质的结合量减少90%[3] |
| 体内研究 (In Vivo) |
将与已形成肿瘤相匹配的小鼠队列随机每天接受载体或 (+)-JQ1 (50 mg/kg) 腹膜内注射。 FDG-PET 成像用于在治疗后四天和随机分组之前评估小鼠。当施用 (+)-JQ1 时,FDG 摄取显着减少。肿瘤体积的评估表明,JQ1 治疗抑制了肿瘤生长。 CD1 小鼠用于口服和静脉给药后 (+)-JQ1 的药代动力学研究。静脉注射(5 mg/kg)后(+)-JQ1 平均血浆浓度的时间曲线。静脉注射(+)-JQ1的半衰期(T1/2)约为1小时,其药代动力学特征显示出良好的药物暴露(AUC=2090 hr*ng/mL)。口服剂量(10 mg/kg)后,创建了 (+)-JQ1 的平均血浆浓度-时间曲线。口服(+)-JQ1药代动力学参数显示出良好的药物暴露(AUC=2090 hr*ng/mL)、血浆峰浓度(Cmax=1180 ng/mL)和口服生物利用度(F=49%)[1]。
1. AML异种移植瘤生长抑制:荷MOLM-13皮下异种移植瘤的裸鼠(肿瘤体积≈100 mm³,n=6/组),经(+)-JQ1(50 mg/kg,口服灌胃,每日1次,21天)或溶媒(5% DMSO+20% Cremophor EL+75%生理盐水)处理。第21天,处理组平均肿瘤体积≈210 mm³,溶媒组≈950 mm³,肿瘤生长抑制率(TGI)≈78%。肿瘤组织中MYC mRNA下降2.8倍,Western blot显示BRD4蛋白减少65%[1] 2. 可逆性男性避孕作用:8周龄雄性C57BL/6小鼠(n=8/组)经(+)-JQ1(25 mg/kg,腹腔注射,每日1次,28天)或溶媒(10% DMSO+90%生理盐水)处理。处理后,附睾精子数量从1.2×10⁷个/附睾降至0.3×10⁷个/附睾,精子活力从65%降至12%。停药4周后,精子数量和活力恢复至正常水平的85%,且无睾丸组织病理学损伤(HE染色)及血清睾酮水平变化[3] |
| 酶活实验 |
乙酰组蛋白结合测定。[1]
如前所述51进行分析,对制造商的方案(PerkinElmer,USA)稍作修改。将所有试剂在补充有0.05%CHAPS的50mM HEPES、100mM NaCl、0.1%BSA、pH 7.4中稀释,并在加入平板之前使其平衡至室温。在150–0μM的范围内制备配体的24点1:2系列稀释液,并将4μl转移到低体积384孔板中,然后加入4μl His标记蛋白(BRD4(1),250 nM,BRD4(2)和CREBBP,2000 nM)。将板密封并在室温下孵育30分钟,然后将4μl等摩尔浓度的生物素化肽添加到蛋白质中[BRD4(1)和BRD4(2)的肽:H4K5acK8acK12acK16ac,HSGRGK(Ac)GGK(Ac。在弱光条件下加入4μl链霉亲和素包被的供体珠(25μg/ml)和4μl镍螯合受体珠(25µg/ml)之前,将板密封并再孵育30分钟。将板箔密封以避光,在室温下孵育60分钟,并使用AlphaScreen 680激发/570发射滤光片组在PHERAstar FS板读取器上读取。对照相应的DMSO对照标准化后,在Prism 5(GraphPad Software,USA)中计算IC50值,并作为20μl反应体积中化合物的最终浓度给出。 1. BRD4结合SPR实验:采用胺偶联法将重组人BRD4 BD1(10 μg/mL)固定于CM5传感器芯片。(+)-JQ1 用运行缓冲液(10 mM HEPES pH7.4、150 mM NaCl、0.05% Tween-20)稀释为0.01–1 μM,以30 μL/min流速注入芯片,记录结合-解离曲线,计算平衡解离常数(Kd)≈0.05 μM[1] 2. ITC结合热力学实验:25°C条件下,将50 μM (+)-JQ1 逐滴加入5 μM BRD4 BD1溶液(缓冲液:20 mM Tris-HCl pH7.5、150 mM NaCl),记录热功率变化。推导结合参数:焓变(ΔH)≈-25 kJ/mol,结合常数(Ka)≈2×10⁷ M⁻¹[1] 3. BRD4抑制HTRF实验:将BRD4 BD1(20 nM)、生物素化组蛋白H4K5ac肽(10 nM)与(+)-JQ1(0.001–10 μM)在反应缓冲液中孵育1小时,加入链霉亲和素-铕与抗BRD4抗体-别藻蓝蛋白,检测665 nm/620 nm荧光比,计算BRD4 BD1的IC₅₀≈0.07 μM[1] |
| 细胞实验 |
细胞增殖测定。[1]
将细胞接种到白色384孔微量滴定板(Nunc)中,在总体积为50μl的培养基中,每孔500个细胞。797、TT和TE10细胞生长在含有1%青霉素/链霉素和10%FBS的DMEM中。Per403细胞生长在含1%青霉素/链霉菌和20%FBS的DMEM中。患者来源的NMC 11060细胞生长在具有10%FBS和1%青霉素/链球菌素的RPMI中。通过机器人销钉转移(PerkinElmer JANUS配备有V&P Scientific 100 nl销钉工具)将化合物递送至微量滴定分析板。在37ºC下孵育48小时后,裂解细胞,并使用商业增殖测定法评估孔的总ATP含量。根据剂量分析重复测量,并通过逻辑回归计算IC50的估计值。[1] 细胞生长测定。[1] 将细胞以每孔1.5×104个细胞的浓度接种在6孔组织培养皿中。细胞在2ml DMEM(797)或RPMI(11060)中生长,所述DMEM或RPMI含有10%胎牛血清、1%青霉素/链霉素和250nM(+)-JQ1或等效体积的DMSO(0.025%)。每天更换每口井中一半的介质。在第0天、第1天、第3天、第7天和第10天,将分配给每个时间点的细胞培养皿进行胰蛋白酶消化,以1:1的比例与0.4%台盼蓝混合,并使用Countess自动细胞计数器计数。 1. MOLM-13细胞MTT抗增殖实验:MOLM-13细胞以5×10³个/孔接种96孔板,用含10% FBS的RPMI 1640培养基过夜培养。加入0.01–1 μM (+)-JQ1,37°C、5% CO₂孵育72小时。每孔加MTT试剂(5 mg/mL,10 μL)孵育4小时,加二甲亚砜(100 μL/孔)溶解甲臜,检测570 nm吸光度,非线性回归计算IC₅₀[1] 2. MYC启动子结合ChIP-qPCR实验:MOLM-13细胞经0.5 μM (+)-JQ1 处理24小时后,用1%甲醛交联,超声破碎染色质。抗BRD4抗体免疫沉淀后,用MYC启动子特异性引物进行qPCR,以Input DNA为参照定量BRD4结合量[1] 3. HeLa细胞自噬检测:HeLa细胞经0.1–1 μM (+)-JQ1 处理48小时。Western blot实验:RIPA裂解细胞,检测LC3B、p62、LAMP1蛋白;免疫荧光实验:4%多聚甲醛固定细胞,抗LC3B抗体(荧光二抗标记)染色,共聚焦显微镜计数LC3B斑点[2] 4. 精原细胞分化实验:从小鼠睾丸分离原代精原细胞,接种6孔板,加入0.05–0.5 μM (+)-JQ1 培养72小时。细胞固定后,抗SYCP3抗体(减数分裂标志物)与DAPI(核染色)染色,荧光显微镜计数SYCP3阳性细胞比例[3] |
| 动物实验 |
1. 溶于 5% 葡萄糖溶液;50 mg/kg;腹腔注射;Nature. 2010 年 12 月 23 日;468(7327):1067-73;
2. 溶于 10% DMSO 和 90% 的 10% 2-羟丙基-β-环糊精溶液;Leukemia. 2017 年 10 月;31(10):2037-2047; 3. 溶于 1% DMSO+5% 葡萄糖+ddH2O;Cell. 2018 年 9 月 20 日;175(1):186-199.e19; 4. 溶于 20% 羟丙基-β-环糊精、5% DMSO、0.2% Tween-80 的生理盐水中; Mol Cancer Ther. 2016 年 6 月;15(6):1217-26.; 5. 溶于 1:1 丙二醇:水混合溶液中;J Biol Chem. 2016 年 11 月 4 日;291(45):23756-23768.; 6. 溶于 5% DMSO 和 10% 2-羟丙基-β-环糊精混合溶液中; Cancer Lett. 2017 年 8 月 28 日;402:100-109。异种移植疗效研究。[1] 将 NMC 797 细胞 (107) 悬浮于 30% Matrigel(BD Biosciences)中,注射到 6 周龄雌性 NCr 裸鼠的侧腹部,建立 NMC 797 异种移植模型。注射 12 天后,将肿瘤可测量的裸鼠分为若干组,分别接受 JQ1(50 mg kg-1,腹腔注射)或载体(5% DMSO 和 5% 葡萄糖溶液)治疗。在FDG-PET研究中,已建立肿瘤(最大线性尺寸约为1 cm)的小鼠在注射250 μCi FDG 1小时后进行基线CT/PET成像(预处理)。随后,小鼠接受腹腔注射,每日一次,每次50 mg/kg的消旋JQ1,连续四天。在第四次注射JQ1或载体后两小时,小鼠再次进行FDG-PET成像(后处理)。整个肿瘤体积内的积分信号以每克注射剂量的百分比(%ID/gm)表示。肿瘤固定于10%中性缓冲福尔马林溶液中进行组织病理学分析。在肿瘤卡尺研究中,JQ1治疗组(n = 8)和载体组(n = 7)在治疗开始时肿瘤的平均大小相似(分别为63.8 ± 17.1 mm³和73.6 ± 14.4 mm³)。每日观察动物的临床症状。肿瘤大小通过游标卡尺测量,体积使用公式 Vol = 0.5 x L x W² 计算。治疗 2 周后,所有小鼠均被实施安乐死,肿瘤固定于 10% 福尔马林溶液中进行组织病理学检查。肿瘤体积的统计学意义采用双侧 Student's t 检验进行计算。[1] 原发性 NMC 异种移植研究。[1] 通过将原代细胞(10⁷ 个细胞溶于 100 μl 30% Matrigel 和 70% PBS 中)注射到 NCr 裸鼠体内,建立了 NMC 原发性异种移植模型。这些原代细胞取自经 Dana-Farber 癌症研究所和 Brigham & Women's 医院伦理委员会批准并获得患者知情同意的恶性胸腔积液。如上所述,四只已形成肿瘤(最大线性尺寸约为1 cm)的小鼠在注射250 μCi FDG 1小时后进行基线CT/PET成像(预处理)。随后,小鼠接受腹腔注射(+)-JQ1,每日一次,每次50 mg/kg,连续四天。每日观察动物的临床症状。在第四次注射(+)-JQ1或载体后两小时,小鼠再次进行FDG-PET成像(后处理)。整个肿瘤体积内的积分信号以每克注射剂量的百分比(%ID/gm)表示。肿瘤固定于10%中性缓冲福尔马林溶液中进行组织病理学分析。研究结束时,所有小鼠均被实施安乐死,肿瘤固定于10%福尔马林溶液中进行组织病理学检查。使用NMC Per403和11060异种移植瘤进行30天生存研究,方法如上所述。在这些研究中,(+)-JQ1以50 mg/kg的剂量每日腹腔注射给药。治疗开始时,(+)-JQ1治疗组(n = 10)和载体组(n = 10)的肿瘤平均大小相似。每日观察动物的临床症状。使用游标卡尺测量肿瘤大小,并使用公式Vol = 0.5×L×W²计算肿瘤体积。采用双侧Student t检验计算肿瘤体积的统计学意义。使用Log-rank(Mantel-Cox)检验进行比较生存分析,并将数据以Kaplan-Meier曲线图的形式呈现,并标注统计学意义(p值)。所有动物实验均已获得丹娜-法伯癌症研究所 (DFCI) 的动物实验伦理委员会 (IACUC) 批准。 小鼠药代动力学研究。[1] 雄性 CD1 小鼠(24-29 克)分别接受单次剂量的 (+)-JQ1 治疗,尾静脉注射剂量为 5 mg/kg,灌胃剂量为 10 mg/kg。在异氟烷麻醉下,于预设时间间隔(0.033、0.083、0.25、0.5、1、2、4、5、8 和 24 小时)通过眼眶后静脉穿刺采集约 150 μl 血液至 EDTA 抗凝管中。每个时间点分析三只动物。血液样本置于冰上,并在采样后 15 分钟内离心(2000 xg,4℃,5 分钟)以获得血浆样本。血浆样本储存在约-70℃直至进行分析。研究期间,小鼠可自由摄取食物和水。化合物配制成10% DMSO和10% HP-β-CD溶液,用于静脉注射。药代动力学研究和药理学分析方法的开发由ChemPartner(中国上海)完成。数据由JEB使用Microsoft Excel和GraphPad Prism 5.02进行分析。 使用的体内制剂(已报道): 1. MOLM-13 AML异种移植模型:将5×10⁶个MOLM-13细胞(悬浮于0.2 mL PBS:Matrigel = 1:1混合液中)皮下注射到雌性裸鼠(6-8周龄,18-22 g)右侧腹部。当肿瘤体积达到约 100 mm³ 时,将小鼠随机分为两组(每组 n=6): - 载体组:灌胃给予 0.2 mL 5% DMSO + 20% Cremophor EL + 75% 生理盐水,每日一次,持续 21 天; - (+)-JQ1 组:灌胃给予 50 mg/kg (+)-JQ1(溶于上述载体中,配制成 250 mg/mL 的溶液),0.2 mL,每日一次,持续 21 天。每 3 天测量一次肿瘤体积(长 × 宽² / 2)和体重。第 22 天处死小鼠,收集肿瘤组织进行 mRNA 和蛋白质分析 [1] 2.雄性避孕小鼠模型:将8周龄、体重22-25克的C57BL/6雄性小鼠随机分为两组(每组n=8): - 溶剂组:腹腔注射0.1 mL 10% DMSO + 90%生理盐水,每日一次,连续28天; - (+)-JQ1组:腹腔注射0.1 mL 25 mg/kg (+)-JQ1(溶于上述溶剂中,配制成250 mg/mL),每日一次,连续28天。 治疗结束后,收集附睾进行精子计数和活力测定。睾丸组织固定后进行HE染色。停药4周后,再次评估精子参数[3] |
| 药代性质 (ADME/PK) |
溴结构域抑制剂 (+)-JQ1 是一种经过高度验证的化学探针;然而,其体内药代动力学性质较差。
这项小型研究中小鼠个体间的显著差异可能会使人们对该药代动力学分析所得平均差异的显著性产生疑问。鉴于我们的主要目标是评估氘代对 (+)-JQ1 清除率的相对影响,而非确定 (+)-JQ1 和 (+)-JQ1-D 的绝对药代动力学参数,我们检测了 (+)-JQ1 及其主要代谢物 M19 的同位素异构体比例,以寻找氘代作用的证据。在雄性和雌性小鼠中,(+)-JQ1/(+)-JQ1-D 比值在所考察的时间进程中持续下降(图 7),表明 (+)-JQ1 的清除速度比 (+)-JQ1-D 更快。[https://pmc.ncbi.nlm.nih.gov/articles/PMC10788937/] 对腹腔注射 30 mg/kg JQ1 的非肿瘤小鼠进行了血浆药代动力学研究。给药后 6 小时采集的所有 JQ1 浓度均低于定量限 (LOQ)。采用线性单室模型可以很好地拟合 JQ1 的总血浆浓度-时间数据,该模型假设腹腔吸收是即时且完全的(图 6)。模型参数估计值见表 7。有限采样模型确定了脑微透析研究期间血浆采样的以下有效时间点:给药后 15 分钟、1 小时和 6 小时。[https://pmc.ncbi.nlm.nih.gov/articles/PMC8384680/] 1. 口服生物利用度:雄性 SD 大鼠(250–300 克,每个时间点 n=3)分别通过灌胃(50 毫克/千克)或静脉注射(10 毫克/千克)给予 (+)-JQ1。口服生物利用度计算为~35%(基于AUC₀₋₂₄ₕ:口服≈15 μM·h;静脉注射≈43 μM·h)[1] 2. 血浆药代动力学:大鼠口服给药(50 mg/kg)后,关键参数为:Cₘₐₓ ≈ 2.8 μM(Tₘₐₓ = 1.5 h),末端半衰期(t₁/₂)≈ 3.2 h,清除率(CL)≈ 18 mL/kg/min。静脉注射(10 mg/kg)后:Cₘₐₓ ≈ 12 μM,t₁/₂ ≈ 2.8 h,CL ≈ 15 mL/kg/min [1] 3. 组织分布:大鼠口服 (+)-JQ1 (50 mg/kg) 1.5 h 后,LC-MS/MS 检测到的浓度:肝脏 ≈ 4.5 μM,肾脏 ≈ 3.2 μM,MOLM-13 异种移植瘤 ≈ 3.8 μM,脑 ≈ 0.3 μM(血脑屏障穿透性低)[1] |
| 毒性/毒理 (Toxicokinetics/TK) |
1. 大鼠亚慢性毒性:口服(+)-JQ1(50 mg/kg,21天)的大鼠体重变化小于对照组(<5%)。血清ALT/AST约为对照组的1.1倍(在正常范围内),肌酐水平正常。外周血白细胞计数约为对照组的0.9倍(无显著差异)[1]
2. 小鼠睾丸安全性:腹腔注射(+)-JQ1(25 mg/kg,28天)的小鼠生精小管结构完整(HE染色),无炎症或坏死。血清睾酮水平约为载体的0.9倍(无显著差异)[3] 3. 血浆蛋白结合率:(+)-JQ1 (1 μM) 在人血浆中的血浆蛋白结合率约为85%(采用30 kDa截留分子量的超滤膜和LC-MS/MS测定)[1] |
| 参考文献 | |
| 其他信息 |
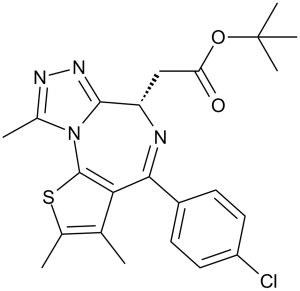
JQ1 属于噻吩并三唑二氮杂卓类化合物,是 [(6S)-4-(4-氯苯基)-2,3,9-三甲基-6H-噻吩并[3,2-f][1,2,4]三唑并[4,3-a][1,4]二氮杂卓-6-基]乙酸的叔丁酯。它是一种溴结构域蛋白4抑制剂,具有抗癌和心脏保护作用。JQ1 可作为溴结构域蛋白4抑制剂、心脏保护剂、抗肿瘤剂、抗炎剂、血管生成抑制剂、细胞凋亡诱导剂和铁死亡诱导剂发挥作用。它是一种噻吩并三唑二氮杂卓类化合物、有机氯化合物、羧酸酯和叔丁酯。
表观遗传蛋白是配体发现中备受关注的靶点。迄今为止,成功的研究仅限于染色质修饰酶,即所谓的表观遗传“写入器”和“擦除器”。尚未有报道有效的组蛋白结合模块抑制剂。本文报道了一种细胞渗透性小分子(JQ1),它能与乙酰赖氨酸识别基序(溴结构域)竞争性结合。通过与溴结构域和末端外结构域(BET)家族成员BRD4的共晶体结构分析,揭示了JQ1与乙酰赖氨酸结合腔具有极佳的形状互补性,从而解释了其对特定人类溴结构域亚群的高效性和特异性。在一种基因明确的、无法治愈的人类鳞状细胞癌亚型中观察到了BRD4的复发性易位。 JQ1 的竞争性结合可将 BRD4 融合癌蛋白从染色质上置换下来,从而诱导鳞状分化,并在 BRD4 依赖性细胞系和患者来源的异种移植模型中产生特异性的抗增殖作用。这些数据为靶向表观遗传“读取器”的蛋白质-蛋白质相互作用提供了概念验证,并为更广泛地开发溴结构域家族的化学探针提供了一个多功能的化学支架。[1] 自噬是一种膜转运过程,它指导溶酶体降解细胞质物质。该过程促进细胞的完整性,虽然自噬的核心机制已被阐明,但促进和维持自噬的机制尚不完全清楚。本文报道,表观遗传读取器 BRD4 和甲基转移酶 G9a 抑制一个不依赖于 TFEB/TFE3/MITF 的转录程序,该程序促进自噬和溶酶体的生物合成。我们发现,BRD4 敲低在某些(但并非所有)情况下均可在体外和体内诱导自噬。在饥饿条件下,涉及 AMPK 和组蛋白去乙酰化酶 SIRT1 的信号级联反应会使染色质结合的 BRD4 解离,从而激活自噬基因并促进细胞存活。重要的是,该过程独立于 BRD4 的促生长特性,且与其存在相互拮抗的关系,并受到 NUT 中线癌驱动因子 BRD4-NUT 的强效抑制。因此,这些发现揭示了一种独特的、选择性的自噬调控机制。[2] 男性避孕的药物治疗方法一直是医学领域长期存在的挑战。为了实现这一目标,我们研究了一种选择性小分子抑制剂(JQ1)对表观遗传阅读蛋白溴结构域和末端外结构域(BET)亚家族的影响。本文报道了一种强效抑制剂,用于抑制睾丸特异性成员BRDT,BRDT在精子发生过程中染色质重塑中起着至关重要的作用。生物化学和晶体学研究证实,JQ1占据BRDT乙酰赖氨酸结合口袋可阻止乙酰化组蛋白H4的识别。用JQ1治疗小鼠可减少生精小管面积、睾丸大小以及精子数量和活力,但不影响激素水平。尽管经JQ1处理的雄性小鼠交配正常,但JQ1在精母细胞和圆形精子细胞阶段的抑制作用可导致完全且可逆的避孕效果。这些数据表明,JQ1是一种新型避孕药,它能够穿过血睾屏障并在精子发生过程中抑制溴结构域活性,为靶向雄性生殖细胞的避孕提供了一种先导化合物。[3] BET抑制机制:(+)-JQ1与BET溴结构域(例如BRD4)的乙酰赖氨酸结合口袋竞争性结合,阻止BET蛋白与染色质结合。这会阻断BET依赖性癌基因(例如MYC)的转录,而这些癌基因驱动多种癌症的增殖[1]。 2. 自噬激活机制:BRD4通过与自噬相关基因(例如ATG5)的启动子结合并抑制其转录来负调控自噬。(+)-JQ1抑制BRD4,解除这种抑制并激活自噬,从而增强癌细胞对营养匮乏的敏感性[2]。 3. 男性避孕潜力:BRDT对精子发生至关重要(仅在减数分裂期间的睾丸生殖细胞中表达)。 (+)-JQ1抑制BRD4,阻断精原细胞分化为精母细胞,导致可逆性少弱精子症。它避免了激素副作用(与传统避孕药不同),使其成为一种非激素男性避孕方法的候选药物[3] |
| 分子式 |
C23H25CLN4O2S
|
|
|---|---|---|
| 分子量 |
456.99
|
|
| 精确质量 |
456.138
|
|
| 元素分析 |
C, 60.45; H, 5.51; Cl, 7.76; N, 12.26; O, 7.00; S, 7.02
|
|
| CAS号 |
1268524-70-4
|
|
| 相关CAS号 |
(R)-(-)-JQ1 Enantiomer;1268524-71-5;JQ-1 (carboxylic acid);202592-23-2
|
|
| PubChem CID |
46907787
|
|
| 外观&性状 |
White to yellow solid
|
|
| 密度 |
1.3±0.1 g/cm3
|
|
| 沸点 |
610.4±65.0 °C at 760 mmHg
|
|
| 闪点 |
322.9±34.3 °C
|
|
| 蒸汽压 |
0.0±1.7 mmHg at 25°C
|
|
| 折射率 |
1.657
|
|
| LogP |
4.49
|
|
| tPSA |
97.61
|
|
| 氢键供体(HBD)数目 |
0
|
|
| 氢键受体(HBA)数目 |
6
|
|
| 可旋转键数目(RBC) |
5
|
|
| 重原子数目 |
31
|
|
| 分子复杂度/Complexity |
706
|
|
| 定义原子立体中心数目 |
1
|
|
| SMILES |
ClC1C([H])=C([H])C(=C([H])C=1[H])C1C2C(C([H])([H])[H])=C(C([H])([H])[H])SC=2N2C(C([H])([H])[H])=NN=C2[C@]([H])(C([H])([H])C(=O)OC(C([H])([H])[H])(C([H])([H])[H])C([H])([H])[H])N=1
|
|
| InChi Key |
DNVXATUJJDPFDM-KRWDZBQOSA-N
|
|
| InChi Code |
InChI=1S/C23H25ClN4O2S/c1-12-13(2)31-22-19(12)20(15-7-9-16(24)10-8-15)25-17(11-18(29)30-23(4,5)6)21-27-26-14(3)28(21)22/h7-10,17H,11H2,1-6H3/t17-/m0/s1
|
|
| 化学名 |
|
|
| 别名 |
|
|
| HS Tariff Code |
2934.99.9001
|
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
|
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
|
|||
|---|---|---|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: ≥ 2.5 mg/mL (5.47 mM) (饱和度未知) in 10% DMSO + 40% PEG300 + 5% Tween80 + 45% Saline (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。
例如,若需制备1 mL的工作液,可将100 μL 25.0 mg/mL澄清DMSO储备液加入到400 μL PEG300中,混匀;然后向上述溶液中加入50 μL Tween-80,混匀;加入450 μL生理盐水定容至1 mL。 *生理盐水的制备:将 0.9 g 氯化钠溶解在 100 mL ddH₂O中,得到澄清溶液。 配方 2 中的溶解度: ≥ 2.5 mg/mL (5.47 mM) (饱和度未知) in 10% DMSO + 90% (20% SBE-β-CD in Saline) (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 例如,若需制备1 mL的工作液,可将 100 μL 25.0 mg/mL澄清DMSO储备液加入900 μL 20% SBE-β-CD生理盐水溶液中,混匀。 *20% SBE-β-CD 生理盐水溶液的制备(4°C,1 周):将 2 g SBE-β-CD 溶解于 10 mL 生理盐水中,得到澄清溶液。 View More
配方 3 中的溶解度: ≥ 2.5 mg/mL (5.47 mM) (饱和度未知) in 10% DMSO + 90% Corn Oil (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液。 配方 4 中的溶解度: 2% DMSO+30% PEG 300+5% Tween 80+ddH2O:5mg/mL 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.1882 mL | 10.9412 mL | 21.8823 mL | |
| 5 mM | 0.4376 mL | 2.1882 mL | 4.3765 mL | |
| 10 mM | 0.2188 mL | 1.0941 mL | 2.1882 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
Leukemia and lymphoma cell lines are broadly sensitive to BET-bromodomain inhibition.Proc Natl Acad Sci U S A.2011 Oct 4;108(40):16669-74. |
Gene expression profiling of LP-1 and Raji cells treated with active or inactive BET inhibitors.Proc Natl Acad Sci U S A.2011 Oct 4;108(40):16669-74. |
Small molecule BET-bromodomain inhibition suppressesMYCtranscription.Proc Natl Acad Sci U S A.2011 Oct 4;108(40):16669-74. |
MYC reconstitution significantly protects cells from BET-mediated effects.Proc Natl Acad Sci U S A.2011 Oct 4;108(40):16669-74. |
BET-bromodomain inhibition decreases tumor load in vivo.Proc Natl Acad Sci U S A.2011 Oct 4;108(40):16669-74. |
Integrated genomic rationale for BET bromodomains as therapeutic targets in MM.Cell.2011 Sep 16;146(6):904-17. |
Inhibition of Myc-dependent transcription by theJQ1BET bromodomain inhibitor.Cell.2011 Sep 16;146(6):904-17. |
BET inhibition suppressesMYCtranscription in MM.Cell.2011 Sep 16;146(6):904-17. |
Regulation ofMYCtranscription by BET bromodomains.Cell.2011 Sep 16;146(6):904-17. |
Anti-myeloma activity ofJQ1in vitro.Cell.2011 Sep 16;146(6):904-17. |
JQ1induces cell cycle arrest and cellular senescence in MM cells.Cell.2011 Sep 16;146(6):904-17. |
Translational implications of BET bromodomain inhibition in MM.Cell.2011 Sep 16;146(6):904-17. |
|
-JQ1") |
-JQ1") |
 RWT9996
RWT9996
 Penipanoid C
Penipanoid C
 (Rac)-EBET-1055
(Rac)-EBET-1055
 SGC-BRDVIII-NC
SGC-BRDVIII-NC
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1
































 COA
COA
-JQ1")
-JQ1")
-JQ1")
-JQ1")
-JQ1")
-JQ1")
-JQ1")
-JQ1")
-JQ1")
-JQ1")
-JQ1")
-JQ1")
-JQ1")
 463611831
463611831
