| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 10 mM * 1 mL in DMSO |
|
||
| 1mg |
|
||
| 2mg |
|
||
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| Other Sizes |
|
| 靶点 |
NMDA receptor (Kd = 1.4 μM); AP-5 acts as a competitive antagonist of the N-methyl-D-aspartate (NMDA) receptor, specifically targeting the glutamate binding site on the NMDA receptor. The compound targets the Glutamate receptor ionotropic NMDA 1 (NMDAR1) subunit. As a competitive antagonist, AP-5 blocks the receptor by competing with the endogenous agonist glutamate for binding at the orthosteric site, thereby preventing receptor activation and the subsequent influx of calcium ions through the receptor channel. No specific IC₅₀ or Kᵢ values for AP-5 at the NMDA receptor were provided in the available literature.
|
|---|---|
| 体外研究 (In Vitro) |
d -对映体是NMDA谷氨酸受体(受体,n -甲基- d -天冬氨酸)的有效特异性拮抗剂。L型对NMDA受体无活性,但可能影响AP4(2-氨基-4-磷酸丁酸酯;兴奋性氨基酸受体。
|
| 体内研究 (In Vivo) |
D-AP5是一种NMDA 受体拮抗剂。长期脑室内 D-AP5 输注会以平行的剂量依赖性方式对体内的长时程增强 (LTP) 和空间学习产生不利影响。当空间学习被阻止时,D-AP5 的大脑浓度不会导致任何可检测到的感觉障碍 [2]。在试验期间,游泳速度的逐渐降低与 D-AP5 的输注有关。当受 D-AP5 影响的动物无法学习时,它们在空间任务中会出现感觉运动异常,并且随着时间的推移会变得更糟。水迷宫的延迟匹配位置方案揭示了用 D-AP5 治疗的大鼠存在延迟依赖性空间记忆缺陷 [3]。
本系列实验研究了NMDA受体拮抗剂d -2-氨基-5-磷酸戊酸酯(D-AP5)是否能在与体内海马长期增强(LTP)损伤相当的剂量范围内诱导空间学习损伤。利用微透析技术估计海马D-AP5的细胞外浓度,以比较这些损伤是否发生在与体外海马切片中损害LTP所需的浓度相似的浓度。将D-AP5以一定浓度(0 ~ 50 mM)通过渗透微型泵长期注入大鼠侧脑室。他们首先接受的训练是在一个开阔的水迷宫任务中找到并逃到一个隐藏的平台上。行为学习结束后,他们被聚氨酯麻醉,并试图唤醒和监测海马LTP。采用微透析法提取海马D-AP5细胞外样本,最后处死动物,解剖组织样本。采用荧光高效液相色谱法分析微透析和组织样品中D-AP5的含量。结果表明,首先,D-AP5对空间学习的损害呈剂量线性依赖关系,与体内海马LTP的损害高度相关。没有观察到D-AP5浓度在不影响学习的情况下阻断LTP。其次,微透析估计表明,在某些假设下,D-AP5在细胞外浓度下导致这些损伤,与体外损害LTP的浓度相当。第三,全组织和微透析样品的比较显示,浓度比约为30:1,表明97%的脑内D-AP5是透析探针无法接近的。研究发现,输注20 mM EGTA可导致透析灌注液中D-AP5增加7倍,这表明至少部分无法进入的D-AP5被钙依赖机制捕获。进一步的两项行为控制研究表明,D-AP5诱导的空间学习障碍不太可能继发于药物诱导的运动障碍,并且D-AP5浓度刚好足以完全阻断海马LTP的组的表现与伊博滕酸诱导的双侧海马病变组的表现在统计学上没有区别。综上所述,这些发现为NMDA受体的激活对于某些类型的学习是必要的这一假设提供了支持。[2] |
| 酶活实验 |
在现有文献中,AP-5的NMDA受体拮抗特性主要通过功能性实验而非直接结合实验来表征。AP-5被描述为一种竞争性NMDA受体拮抗剂,意味着它与内源性激动剂谷氨酸竞争结合NMDA受体的正构(谷氨酸识别)位点。该化合物对NMDA受体相对于其他谷氨酸受体亚型(如AMPA和红藻氨酸受体)的特异性已通过电生理和药理学研究得到充分证实,使其成为区分NMDA受体介导反应与其他谷氨酸能信号传导的标准工具。
|
| 细胞实验 |
现有文献中关于AP-5的详细细胞实验方案有限。在培养的神经元模型中,浓度为100 μM的AP-5可部分阻止谷氨酸诱导的Arc/Arg3.1蛋白水平升高,Arc/Arg3.1是活性依赖性基因表达的标志物。该化合物还降低了NMDA诱导的Arc/Arg3.1上调,证实了其在细胞模型中阻断NMDA受体介导信号传导的能力。这些实验通常涉及在NMDA或谷氨酸刺激之前或同时用AP-5处理培养的神经元,随后提取蛋白并通过Western blot分析测量Arc/Arg3.1水平。
|
| 动物实验 |
药物浓度、手术、氨基酸分析和组织学:除最终重复实验外,所有药物和麻醉剂均用去离子水配制,最终重复实验则使用无热原水配制。将等浓度的100 mM D-AP5储备液用100 mM NaOH配制,并冷冻保存。用人工脑脊液(aCSF)稀释至一系列浓度(5、13、20、30、40和50 mM),并加入NaOH以维持pH值为7.4。将等浓度的EGTA储备液用NaOH配制,并用无钙aCSF稀释至20 mM。将异戊烯酸溶解于pH 7.4的磷酸盐缓冲液中,浓度为10 mg/ml。由于D-AP5无法穿过血脑屏障,因此采用渗透泵(Alza 2002型;泵送速率0.5 pl/hr)进行慢性输注,将药物输送至脑部。将动物固定于Kopf立体定位仪上,在三溴乙醇麻醉(0.29 g/kg)下植入渗透泵。沿中线切开,暴露颅骨,将L形不锈钢套管(23号)置入左侧脑室(距颅骨表面AP = -0.9,ML = 1.3,DV = -4.5)。套管通过硅胶管连接至渗透泵,并用牙科丙烯酸树脂和三颗钟表匠螺丝固定。渗透泵置于从切口尾端延伸至肩胛骨的皮下囊袋内。头皮切口采用间断缝合,术后动物恢复2天。手术过程中,在颅骨上标记微透析探针和两个电极的位置。对于异戊烯酸损伤实验,将动物固定于立体定位仪上,并用三溴乙醇麻醉。根据Jarrard(1989)的方法,在双侧26个位点使用异戊烯酸(0.05或0.10 μL)进行损伤,并在行为学测试前恢复2周。使用Varian Vista 5500泵系统、9090自动进样器和5 μm Nucleosil C-18色谱柱(250 mm × 4.6 mm)分析脑组织和透析样品中的D-AP5和氨基酸。采用梯度洗脱法分离氨基酸,洗脱液为磷酸盐缓冲液[缓冲液A:50 mM 和四氢呋喃 (THF) (2.5%);pH 5.121,有机改性剂为甲醇[缓冲液B:和 THF (1.25%)]。梯度洗脱程序如下(泵速为 1 ml/min):[时间 (min), %B] 0, 0; 5, 0; 7, 25; 15, 50; 23, 60; 25.90: 28.110: 32.100: 42.0。柱前用邻苯二甲醛衍生化(L&d&h 等,1915)后,使用 ABS 980 荧光检测器(激发波长 230 pm;发射波长 1398 pm)通过荧光法检测伯氨基酸。在每日分析的开始和结束时,将含有已知量氨基酸和D-AP5的标准品注入色谱柱,以校准每种待测分子组分的保留时间和峰面积。使用基于微型计算机的积分软件包对数据进行积分和定量。实验结束后处死动物,并在冰上取出脑组织。从剂量反应研究中的动物的左右海马中解剖出组织,并冷冻保存(-20°C),用于分析D-AP5和氨基酸含量。将紧邻输注套管的组织保存在福尔马林中,冷冻,切成30 μm厚的冠状切片,并用快甲酚紫染色。这有助于验证套管的位置,并评估其和/或药物输注造成的任何损伤。将海马损伤动物的大脑包埋于蛋黄中,冷冻后切成30 μm厚的水平切片,以评估细胞丢失程度。[2]
为了验证海马N-甲基-D-天冬氨酸(NMDA)受体直接参与海马依赖性学习机制的假设,我们进行了三项实验,以对比NMDA受体拮抗剂引起的学习障碍是由于药物引起的神经病理学和/或感觉运动障碍所致。在实验1中,与注射人工脑脊液(aCSF)的动物相比,接受d-AP5(30 mm)慢性脑室内(icv)输注(0.5 μL/h)的大鼠在14天的药物输注期间进行训练时,其位置学习能力选择性受损,但在线索导航学习能力方面未受影响;但如果在微型泵停止输注11天后进行训练,则两种任务均未受损。 d-AP5 导致空间任务中的感觉运动障碍,但随着动物学习能力的下降,这些障碍逐渐加重。对潜在神经病理学变化的组织学评估显示,无论是在慢性药物输注期间还是之后处死的 d-AP5 处理组大鼠,均未发现异常。在实验 2 中,接受 d-AP5 处理并训练进行空间参考记忆任务的大鼠也表现出空间学习缺陷。该任务涉及两个相同但可见的平台,经选择和验证可最大程度地减少感觉运动障碍。采用高效液相色谱法 (HPLC) 检测了特定脑区中 d-AP5 的存在。在实验 3 中,接受 d-AP5 处理的大鼠在水迷宫延迟匹配位置实验中表现出延迟依赖性的空间记忆缺陷。本文讨论了这些数据与学习机制以及 NMDA 受体拮抗剂对脑功能影响的感觉运动机制之间的关系。我们认为 NMDA 受体机制直接参与空间学习。[3] |
| 药代性质 (ADME/PK) |
现有文献中未描述AP-5的ADME/药代动力学性质(吸收、分布、代谢、排泄、半衰期、口服生物利用度、血浆蛋白结合率)。AP-5主要用作动物研究中脑室内、海马内或腹腔注射给药的研究工具。该化合物具有水溶性,便于用于注射制剂。
|
| 毒性/毒理 (Toxicokinetics/TK) |
现有文献中未报道AP-5的直接毒性数据(如LD₅₀、肝毒性、肾毒性或特定器官毒性)。该化合物仅用于研究目的,不适用于人类治疗用途。在动物研究中,AP-5以多种剂量给药而未报告急性毒性:大鼠腹腔注射剂量高达0.7 mg/kg/天持续7天,脑室内注射剂量高达10 nmol,以及小鼠CA1区内注射剂量高达0.5 μg/小鼠。处理该化合物时应采取标准的实验室安全预防措施,包括使用适当的个人防护装备。
|
| 参考文献 |
|
| 其他信息 |
D-对映体是NMDA谷氨酸受体(受体,N-甲基-D-天冬氨酸)的强效且特异性拮抗剂。L型对NMDA受体无活性,但可能影响AP4(2-氨基-4-膦酸丁酸;APB)兴奋性氨基酸受体。
1 在蛙或幼鼠的离体脊髓标本中,研究了一系列ω-膦酸α-羧酸氨基酸对诱发电活动的抑制作用以及兴奋性氨基酸拮抗特性。 2. 在背根诱发的腹根电位测试中,从2-氨基-5-磷酸戊酸到2-氨基-8-磷酸辛酸的同系物均表现出抑制作用,这与这些物质拮抗N-甲基-D-天冬氨酸诱导的选择性运动神经元去极化的能力相关。3. 2-氨基-5-磷酸戊酸是该系列中最有效的物质,其对N-甲基-D-天冬氨酸反应的拮抗作用的表观KD值为1.4 μM。4. 2-氨基-5-磷酸戊酸的(+)-和(-)-异构体的比较表明,该物质的N-甲基-D-天冬氨酸拮抗活性和神经元抑制作用主要归因于(-)-异构体。 5. 2-氨基-4-膦酸丁酸酯的(-)-型和(+)-型作用不同。该物质的(-)-型对N-甲基-D-天冬氨酸、奎斯奎酸和红藻氨酸诱导的去极化具有相对较弱的非选择性拮抗作用,并且在对诱发电活动进行测试时也表现出类似的弱抑制作用。(+)-型抑制电诱发活动的能力强于(-)-型,但并不拮抗氨基酸兴奋剂的反应。当浓度高于抑制电诱发活动所需的浓度时,(+)-型会引起去极化。这种作用可被2-氨基-5-膦戊酸酯阻断。[1] |
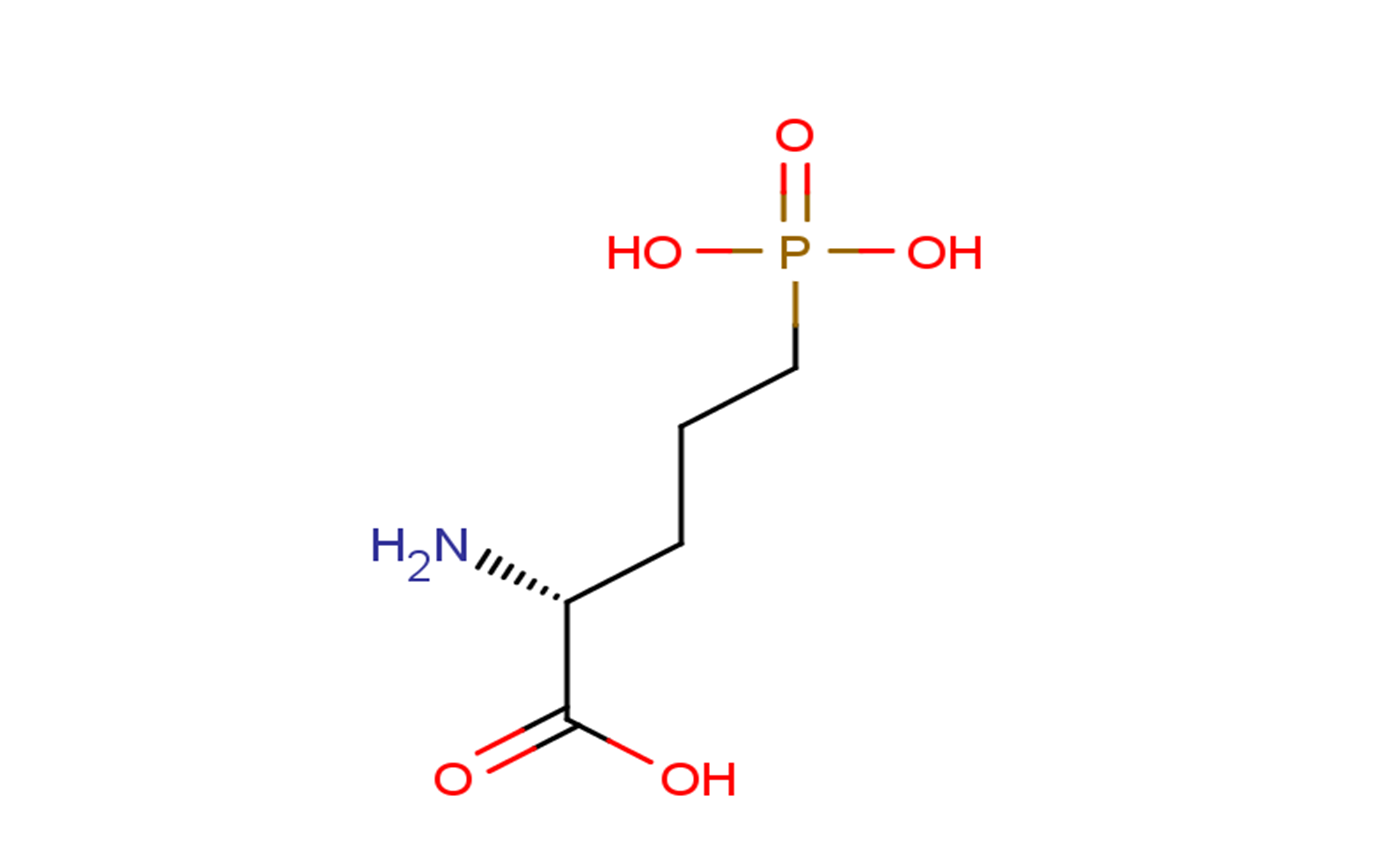
| 分子式 |
C5H10NO5P-2
|
|---|---|
| 分子量 |
195.1104
|
| 精确质量 |
197.045
|
| 元素分析 |
C, 30.47; H, 6.14; N, 7.11; O, 40.58; P, 15.71
|
| CAS号 |
79055-68-8
|
| 相关CAS号 |
DL-AP5;76326-31-3;L-AP5;79055-67-7
|
| PubChem CID |
135342
|
| 外观&性状 |
White to off-white solid powder
|
| 密度 |
1.5±0.1 g/cm3
|
| 沸点 |
482.1±55.0 °C at 760 mmHg
|
| 闪点 |
245.4±31.5 °C
|
| 蒸汽压 |
0.0±2.6 mmHg at 25°C
|
| 折射率 |
1.536
|
| LogP |
-2.32
|
| tPSA |
130.66
|
| 氢键供体(HBD)数目 |
4
|
| 氢键受体(HBA)数目 |
6
|
| 可旋转键数目(RBC) |
5
|
| 重原子数目 |
12
|
| 分子复杂度/Complexity |
200
|
| 定义原子立体中心数目 |
1
|
| SMILES |
C(C[C@H](C(=O)O)N)CP(=O)(O)O
|
| InChi Key |
VOROEQBFPPIACJ-SCSAIBSYSA-N
|
| InChi Code |
InChI=1S/C5H12NO5P/c6-4(5(7)8)2-1-3-12(9,10)11/h4H,1-3,6H2,(H,7,8)(H2,9,10,11)/t4-/m1/s1
|
| 化学名 |
(2R)-2-amino-5-phosphonopentanoic acid
|
| 别名 |
D-AP5; (R)-2-Amino-5-phosphonopentanoic acid; 5-Phosphono-D-norvaline; (2R)-2-amino-5-phosphonopentanoic acid; d-APV; D-Norvaline, 5-phosphono-; D-(-)-2-Amino-5-phosphonopentanoic Acid;
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
H2O : ~27.78 mg/mL (~140.92 mM)
DMSO :< 1 mg/mL Ethanol :< 1 mg/mL |
|---|---|
| 溶解度 (体内实验) |
配方 1 中的溶解度: 100 mg/mL (507.28 mM) in PBS (这些助溶剂从左到右依次添加,逐一添加), 澄清溶液; 超声助溶。
请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 5.1253 mL | 25.6266 mL | 51.2531 mL | |
| 5 mM | 1.0251 mL | 5.1253 mL | 10.2506 mL | |
| 10 mM | 0.5125 mL | 2.5627 mL | 5.1253 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
 肌氨酸
肌氨酸
 NMDAR 拮抗剂1
NMDAR 拮抗剂1
 雷拉地尔
雷拉地尔
 非氨酯
非氨酯
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1
































 COA
COA
 463611831
463611831
