| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg | |||
| 100mg |

| 靶点 |
Fluorescent dye
|
|---|---|
| 体外研究 (In Vitro) |
LysoTracker Red 染色实验流程
1. 工作液配制 • 步骤 1.1:将冻存的 1 mM LysoTracker Red 储存液恢复至室温,瞬时离心使液体沉降于管底。 • 步骤 1.2:使用培养基(如 DMEM、RPMI-1640)或缓冲液(如 PBS)稀释储存液,配制成 50-100 nM 的工作液(具体浓度可根据实验需求调整)。 注意:建议现配现用,避免长时间存放影响染色效果。 2. 细胞染色步骤 2.1 细胞准备 • 悬浮细胞: o 4℃、1000×g 离心 3-5 分钟,弃上清。 o 用 PBS 洗涤 2 次,每次 5 分钟。 • 贴壁细胞: o 弃去培养基,胰酶消化后制成单细胞悬液。 o 4℃、1000×g 离心 3-5 分钟,弃上清。 o 用 PBS 洗涤 2 次,每次 5 分钟。 2.2 染色孵育 • 加入 1 mL LysoTracker Red 工作液,室温避光孵育 5-30 分钟(时间可根据细胞类型优化)。 2.3 洗涤与检测 • 4℃、400×g 离心 3-4 分钟,弃上清。 • PBS 洗涤 2 次,每次 5 分钟。 • 用 无血清培养基或 PBS 重悬细胞,进行 荧光显微镜观察 或 流式细胞术分析。 注:实验过程中建议避光操作,以减少荧光淬灭。 LysoTracker Red染料已被用于显微镜成像酸性球形细胞器(Chikte等人,2014)。暴露于BDE-100的HepG2细胞表现出溶酶体染料的细胞染色增加,其积累与自噬过程中典型的溶酶体酸化成正比(图1)。对照组和BDE-100处理的细胞在定性(图1A)或定量(图1B)上存在差异。与对照组相比,暴露于BDE-100 48小时后,LysoTracker染色更加强烈,这表明溶酶体空泡形成增加。假设BDE-100可以诱导自噬,如上述结果所示,我们的下一步是评估HepG2细胞中自噬关键蛋白质的免疫荧光和蛋白质含量。为此,在0.1-25μM的范围内选择了三种浓度并进行了测定。[2] ε毒素诱导液泡的特性[3] 我们研究了LysoTracker Red进入细胞的情况,这是一种荧光嗜酸探针,在中性pH下穿透细胞膜,并分布在活细胞中的酸性细胞器中,如晚期内体和溶酶体(图5C)。在未经ε毒素处理的对照细胞中,LysoTracker标记的囊泡散布在整个细胞质中。在用ε毒素处理240小时的细胞中 min,溶血素红积聚在液泡中,表明液泡呈酸性。 Rab5和Rab7对ε毒素诱导的空泡化的影响[3] 据报道,Rab7是晚期内体和溶酶体的标志物,在VacA处理的细胞中定位于液泡膜,但Rab5是早期内体的标志物。我们确定了Rab5和Rab7是否分布在ε毒素处理的细胞中的液泡膜上。将过表达绿色荧光蛋白(GFP)-Rab5和GFP-Rab7的MDCK细胞暴露于ε毒素。如图6A、B所示,液泡膜用GFP-Rab7标记,但不用GFP-Rab5标记。这一结果表明,ε毒素引起的液泡起源于晚期内体和溶酶体。为了证实这一点,用野生型和显性阴性Myc-Rab5和Flag-Rab7瞬时转染的MDCK细胞在37℃下与ε毒素一起孵育 240°C 通过直接显微镜观察定量空泡化。图6C显示了用Myc抗体或Flag抗体检测的细胞裂解物的蛋白质印迹。24小时后的表达水平 h对于每种蛋白质的野生型和突变型都是相似的。如图6E所示,显性负Rab5的表达部分减少了毒素引起的空泡化。另一方面,图6F显示,与单独的载体和野生型Rab7相比,Rab7的显性负型强烈抑制空泡化。如图6D所示,Rab5DN和Rab7DN的表达可防止毒素诱导的细胞死亡,并抑制空泡化。 |
| 体内研究 (In Vivo) |
背景:LysoTracker Red(LT)是一种多聚甲醛固定探针,可浓缩到细胞和组织的酸性区室中。细胞死亡后,由于邻近细胞吞噬凋亡体,组织中表达了高水平的溶酶体活性(酸性酶)。LT之前被证明是细胞死亡的指标,其方式类似于其他标准检测(膜联蛋白、末端dUTP缺口末端标记、硫酸尼罗蓝、中性红和吖啶橙)。
方法:使用共聚焦激光扫描显微镜(CLSM)在妊娠第14天服用致畸原5-氟尿嘧啶(5-FU)8小时后测量胎鼠后肢的LT荧光。研究了5-FU的四个剂量水平(0、20、30和40mg/kg)。制备技术包括LT染色、多聚甲醛固定、甲醇脱水、苯甲醇和苯甲酸苄酯清除。经过这种治疗,肢体几乎透明,可以进行CLSM分析。 结果:在正常(对照)后肢发生凋亡的特定区域观察到LT染色。5-FU治疗后,肢体的进展区(PZ)出现了高荧光区域。观察到5-FU治疗的剂量依赖性反应。与对照组相比,用20、30和40mg/kg 5-FU治疗的后肢在高度增殖的PZ内显示出更多的荧光。这些结果表明5-FU暴露与LT摄取之间存在剂量反应关系。 结论:我们发现,三种不同的计算机分析程序可以量化胚胎肢芽中指示高水平荧光的三维体积区域。清除组织的样品制备程序、解决设备变量的CLSM技术以及统计总体分析程序的应用相结合,使厚度约为500微米的整个胎鼠后肢的荧光可视化和定量成为可能。 |
| 细胞实验 |
LysoTracker监测溶酶体分布[2]
由于LysoTracker Red染料保留在细胞颗粒内,因此它们已被用于研究细胞中的自噬程度(Rodriguezenriquez等人,2006,Chikte等人,2014)。对于这些测定,将HepG2细胞接种在六孔板的玻璃盖玻片上,然后用BDE-100或不用BDE-10处理。接下来,根据制造商的说明,用磷酸缓冲盐水(PBS)轻轻洗涤细胞,并在不含酚红的培养基中用100 nM LysoTracker Red加1μM Hoechst 33342孵育约30分钟。然后,借助尼康Eclipse TS100荧光显微镜对细胞进行检查和拍照。比较细胞,计数拍摄细胞中的点状Lysotracker模式事件,并表示为相对于阴性对照的百分比(n=3)。 免疫荧光染色和共聚焦成像[3] 将MDCK细胞铺在聚赖氨酸包被的玻璃底培养皿上,并在37℃下孵育 在5%CO2培养箱中,在胎牛血清/DMEM中过夜。为了研究毒素的内化,将ε毒素与细胞在4℃下孵育 1°C h在胎牛血清/DMEM中。在冷胎牛血清/DMEM中洗涤三次后,将细胞转移到预热至37℃的胎牛血清-DMEM中 °C,在相同温度下孵育4小时 h.用冷NaCl/Pi洗涤四次,并在室温下用4%多聚甲醛固定。对于抗体标记,然后将培养皿在室温下孵育15分钟 50分钟 在NaCl/Pi中的NH4Cl浓度为20 含0.1%Triton的NaCl/Pi中的最小值 X-100在室温下。用含0.02%Triton的NaCl/Pi洗涤后 X-100,将培养皿在室温下孵育1小时 h,用含4%BSA的NaCl/Pi溶液,然后在室温下用含4%BSA/Pi溶液中的第一抗体处理1小时 h.然后用含有0.02%Triton的NaCl/Pi洗涤 X-100,与二抗(Alexa Fluor 488-结合抗兔IgG或Alexa Fluor 568-共轭抗小鼠IgG)在含有4%BSA的NaCl/Pi中在室温下放置1小时 h、 用含0.02%Triton的NaCl/Pi广泛洗涤 X-100,并在尼康下进行分析 A1激光扫描共聚焦显微镜。肌动蛋白丝和核分别用罗丹明-鬼笔环肽和DAPI染色。对于使用LysoTracker RedDND-99的实验,MDCK细胞与LysoTracker RedDND-1999(100 nm)和Hoechst一起孵育 33342 (20 μg·mL−1)在37℃下 30°C 活细胞成像前分钟。所有图像都表示穿过焦平面的单个部分。 |
| 动物实验 |
Animal Treatment and Sample Preparation [1]
Timed-pregnant Sprague-Dawley rats were obtained from Charles River Laboratories (Raleigh, NC). The presence of a copulatory plug was designated GD 0. On GD 14, pregnant rats were injected subcutaneously with 20, 30, or 40 mg/kg of 5-FU or saline (control) and killed 8 h later. The pregnant rats were killed by decapitation, the fetuses were removed, and the hindlimbs were dissected immediately and placed in warm phosphate buffered saline (PBS) before incubation. Groups consisting of four hindlimbs each were incubated in 0.5 ml of this medium at 37°C. Sample Preparation: Staining, Fixation, Dehydration, and Clearing [1] The procedure to stain embryonic tissue with LysoTracker Red/LT was described previously. Briefly, a vial of LysoTracker Red/LT containing 50 μl of a 1 mM solution was added to 10 ml of PBS to make a final concentration of 5 μM. PF (20%) was diluted to 4% with PBS and stored frozen at −20°C (14). Limbs were washed twice with PBS after the 30-min staining period, fixed in 4% PF at 4°C overnight, and processed within 24 h. The fetal tissues were washed twice with PBS to remove the fixative and then dehydrated with methanol (MeOH). The limbs were cleared with BABB (1:2 by volume) to produce a nearly transparent limb bud (14, 23, 24). The limbs were placed first into a 1:1 solution of MeOH and BABB for a few hours and then into 100% BABB. The limbs were then transferred into an instrument shop made a ⅛-in. aluminum slide containing a ½-in. hole and sealed. Laser Scanning Confocal Microscopy [1] The Leica CLSM consisted of a Leica inverted DMIRBE microscope and an Omnichrome laser emitting at three wavelengths (488, 568, and 647 nm). The 568-nm line using a triple-dichroic beam splitter–excited LysoTracker Red/LT dye with a slit between 580 and 630 nm was used to measure the emitted light. To visualize an entire GD 14 rat hindlimb with sufficient fluorescent intensity, a Zeiss 5× objective (0.25 numerical aperture [NA]) was used. A Dell 420 workstation with two 933-MHz processors and 1 gigabyte of random access memory and an Nvida Gforce2 video 32-megabyte board were used for the analysis. Image Acquisition [1] The CLSM was evaluated to ensure it was stable and produced good resolution, field illumination, and stable laser power as described in two recent publications from our laboratory. The limb was optically sectioned into 20 to 35 sections with a constant 20-μm thickness between sections. Each 3D volume data set completely encompassed the entire depth (400–700 μm) of the limb tissue. The 40-mg/kg 5-FU–treated limbs containing the brightest fluorescence was measured first, and all confocal settings were kept constant for the other dose groups. These settings included laser power, photomultiplier (PMT) voltage, PMT offset, frame averaging, and step distances between adjacent sections. The size of each image section was 512 × 512 pixels and occupied a region of 1.9 × 1.9 mm. The fluorescent intensity at each voxel was detected and digitized into TIFF images retaining 8 bits of information, or 256 intensity levels. Thus the file size of a representative 512 × 512 × 30 × 8 volume data set was approximately 8 megabytes. Visualization [1] The following software products were used to analyze the raw data: TCS-SP1, VoxBlast, Image-Pro Plus, and Imaris/Surpass. Image Segmentation [1] Each 3D maximum projection image of limbs was segmented into the following regions: whole limb, apical ectodermal ridge (AER), and progress zone (PZ; Fig. 1). The analysis focused on the highly proliferative region of mesenchymal tissue at the front of the limb paddle, the PZ. This region contains proliferating cells that are very reactive to 5-FU and other chemotherapeutic drugs and accumulate most of the LysoTracker Red/LT stain. The PZ was arbitrarily determined to extend 450 μm from the most distal point of the limb into the interior of the limb from a two-dimensional image. A perpendicular line intersecting this interior endpoint was made, and all regions distal to this line were cropped from the image, as shown in Figure 1. The segmentation process for the AER can be performed manually or with an edge-detection technique to remove it from the quantitative analysis procedures. |
| 参考文献 |
|
| 其他信息 |
DND-99 dye is a BODIPY dye. It has a role as a fluorochrome.
To reduce flammability and meet regulatory requirements, Brominated Flame Retardants (BFRs) are added to a wide variety of consumer products including furniture, textiles, electronics, and construction materials. Exposure to polybrominated phenyl ethers (PBDEs) adversely affects the human health. Bearing in mind that (i) PBDEs are potentially toxic, (ii) the mechanism of PBDE toxicity is unclear, and (iii) the importance of the autophagy to the field of toxicology is overlooked, this study investigates whether an autophagic process is activated in HepG2 cells (human hepatoblastoma cell line) to mediate BDE-100-induced toxicity. HepG2 cells were exposed with BDE-100 at three concentrations (0.1, 5, and 25μM), selected from preliminary toxicity tests, for 24 and 48h. To assess autophagy, immunocytochemistry was performed after exposure of HepG2 cells to BDE-100. Labeling of HepG2 cells with 100nM LysoTracker Red DND-99 aided examination of lysosome distribution. Proteins that are key to the autophagic process (p62 and LC3) were evaluated by western blotting. DNA was isolated and quantified to assess mitochondrial DNA copy number by qPCR on the basis of the number of DNA copies of a mitochondrial encoded gene normalized against a nuclear encoded gene. Conversion of LC3-I to LC3-II increased in HepG2 cells. Pre-addition of 100nM wortmannin decreased the amount of LC3 in the punctuate form and increased nuclear fragmentation (apoptotic feature). HepG2 cells exposed to BDE-100 presented increased staining with the lysosomal dye and had larger LC3 and p62 content after pre-treatment with ammonium chloride. The mitochondrial DNA copy number decreased, which probably constituted an attempt of the cell to manage mitochondrial damage by selective mitochondrial degradation (mitophagy). In conclusion, an autophagic process is activated in HepG2 cells to mediate BDE-100-induced toxicity. [2] The epsilon-toxin of Clostridium perfringens forms a heptamer in the membranes of Madin-Darby canine kidney cells, leading to cell death. Here, we report that it caused the vacuolation of Madin-Darby canine kidney cells. The toxin induced vacuolation in a dose-dependent and time-dependent manner. The monomer of the toxin formed oligomers on lipid rafts in membranes of the cells. Methyl-β-cyclodextrin and poly(ethylene glycol) 4000 inhibited the vacuolation. Epsilon-toxin was internalized into the cells. Confocal microscopy revealed that the internalized toxin was transported from early endosomes (early endosome antigen 1 staining) to late endosomes and lysosomes (lysosomal-associated membrane protein 2 staining) and then distributed to the membranes of vacuoles. Furthermore, the vacuolation was inhibited by bafilomycin A1, a V-type ATPase inhibitor, and colchicine and nocodazole, microtubule-depolymerizing agents. The early endosomal marker green fluorescent protein-Rab5 and early endosome antigen 1 did not localize to vacuolar membranes. In contrast, the vacuolar membranes were specifically stained by the late endosomal and lysosomal marker green fluorescent protein-Rab7 and lysosomal-associated membrane protein 2. The vacuoles in the toxin-treated cells were stained with LysoTracker Red DND-99, a marker for late endosomes and lysosomes. A dominant negative mutant of Rab7 prevented the vacuolization, whereas a mutant form of Rab5 was less effective. These results demonstrate, for the first time, that: (a) oligomers of epsilon-toxin formed in lipid rafts are endocytosed; and (b) the vacuoles originating from late endosomes and lysosomes are formed by an oligomer of epsilon-toxin. [3] Collectively, the experimental data provided here showed that BDE-100 modulates autophagic markers in human hepatoblastoma cells (HepG2). Increased LC3-I levels, augmented LC3-II conversion, appearance of dot-like formation by lysosomes as probed with lysoTracker and LC3 by immunofluorescence, higher p62 levels after blocking of the lysosomal activity, and smaller mitochondrial DNA copy number suggested selective mitochondrial degradation (damaged mitochondria) by autophagy. In addition, results revealed that autophagy exerted a protective effect against BDE-100 toxicity; indeed, pretreatment of the cells with wortmannin induced cell death by apoptosis. [2] We investigated the internalization of epsilon-toxin by endocytosis. When the toxin was incubated with MDCK cells at 37 °C, it colocalized with EEA1 after 30 min, indicating that it reaches the early endosomes. The toxin no longer localized with EEA1 at 60 min. After 60 min, it colocalized with Lamp2, indicating that the toxin moves to the late endosomes and lysosomes. These results demonstrate that epsilon-toxin is endocytosed, and sorted from early endosomes to late endosomes and lysosomes. Epsilon-toxin induces the formation of cytoplasmic vacuoles. The toxin was internalized by cells and distributed in association with vacuolar membranes, in a distribution similar to that of Rab7 and Lamp2, but not EEA1 and Rab5. The abundance of Rab7 and Lamp2 in the membranes of epsilon-toxin-generated vacuoles suggests that the vacuolar membranes are derived from late endosomes and lysosomes. Furthermore,LysoTracker Red accumulated in vacuoles of epsilon-toxin-treated cells. Several Rab proteins have been implicated in membrane trafficking along the endocytic pathway. Rab5 regulates early endocytic trafficking as well as the overall organization of early endosomes [26]. Overexpression of dominant negative mutants of Rab5 partially, although not completely, inhibited the formation of vacuoles by epsilon-toxin. GFP–Rab5 was absent from vacuoles. Rab7 actually participates in late endosome homotypic fusion and the transport between late endosomes and lysosomes [26]. In this study, Rab7 was present on the membranes of vacuoles. Cells expressing the dominant negative forms of Rab7 did not develop vacuoles when exposed to epsilon-toxin. Upstream blockade of membrane transport to late endosomes prevented the vacuoles from developing, as described above. The present study indicates that Rab7 plays an essential role in the development of vacuoles induced by epsilon-toxin, and that efficient membrane flow from the plasma membrane to late endosomes and lysosomes, mediated by early endosomes, is crucial to the vacuolation process. [3] |
| 分子式 |
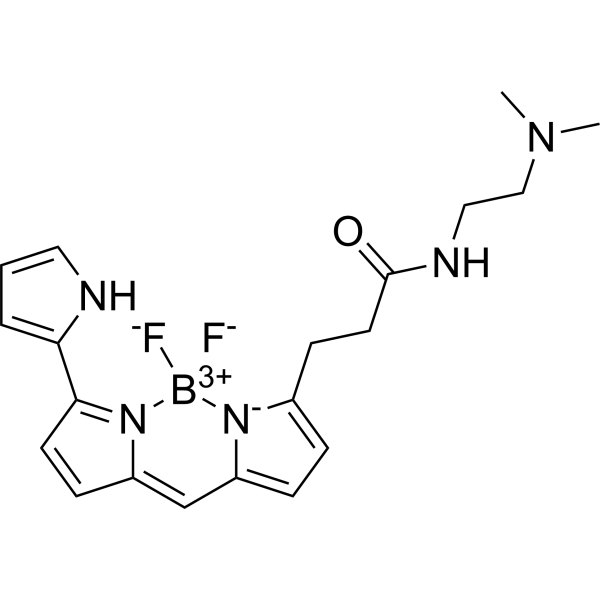
C20H24BN5OF2
|
|---|---|
| 分子量 |
399.245
|
| 精确质量 |
399.204
|
| CAS号 |
231946-72-8
|
| PubChem CID |
15410449
|
| 外观&性状 |
Brown to dark brown solid powder
|
| tPSA |
56.1
|
| 氢键供体(HBD)数目 |
2
|
| 氢键受体(HBA)数目 |
5
|
| 可旋转键数目(RBC) |
7
|
| 重原子数目 |
29
|
| 分子复杂度/Complexity |
751
|
| 定义原子立体中心数目 |
0
|
| SMILES |
[F-][B+3]1([N-]2C(=CC=C2C=C2C=CC(C3NC=CC=3)=N12)CCC(=O)NCCN(C)C)[F-]
|
| InChi Key |
DYYUXMKNXUZBMO-UHFFFAOYSA-N
|
| InChi Code |
InChI=1S/C20H24BF2N5O/c1-26(2)13-12-25-20(29)10-8-15-5-6-16-14-17-7-9-19(18-4-3-11-24-18)28(17)21(22,23)27(15)16/h3-7,9,11,14,24H,8,10,12-13H2,1-2H3,(H,25,29)
|
| 化学名 |
3-[2,2-difluoro-12-(1H-pyrrol-2-yl)-3-aza-1-azonia-2-boranuidatricyclo[7.3.0.03,7]dodeca-1(12),4,6,8,10-pentaen-4-yl]-N-[2-(dimethylamino)ethyl]propanamide
|
| 别名 |
Lysotracker Red; DND-99 dye; 231946-72-8; LysoTracker Red DND-99; CHEBI:52117;
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month 注意: 本产品在运输和储存过程中需避光。 |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO : ~5 mg/mL (~12.52 mM)
|
|---|---|
| 溶解度 (体内实验) |
注意: 如下所列的是一些常用的体内动物实验溶解配方,主要用于溶解难溶或不溶于水的产品(水溶度<1 mg/mL)。 建议您先取少量样品进行尝试,如该配方可行,再根据实验需求增加样品量。
注射用配方
注射用配方1: DMSO : Tween 80: Saline = 10 : 5 : 85 (如: 100 μL DMSO → 50 μL Tween 80 → 850 μL Saline)(IP/IV/IM/SC等) *生理盐水/Saline的制备:将0.9g氯化钠/NaCl溶解在100 mL ddH ₂ O中,得到澄清溶液。 注射用配方 2: DMSO : PEG300 :Tween 80 : Saline = 10 : 40 : 5 : 45 (如: 100 μL DMSO → 400 μL PEG300 → 50 μL Tween 80 → 450 μL Saline) 注射用配方 3: DMSO : Corn oil = 10 : 90 (如: 100 μL DMSO → 900 μL Corn oil) 示例: 以注射用配方 3 (DMSO : Corn oil = 10 : 90) 为例说明, 如果要配制 1 mL 2.5 mg/mL的工作液, 您可以取 100 μL 25 mg/mL 澄清的 DMSO 储备液,加到 900 μL Corn oil/玉米油中, 混合均匀。 View More
注射用配方 4: DMSO : 20% SBE-β-CD in Saline = 10 : 90 [如:100 μL DMSO → 900 μL (20% SBE-β-CD in Saline)] 口服配方
口服配方 1: 悬浮于0.5% CMC Na (羧甲基纤维素钠) 口服配方 2: 悬浮于0.5% Carboxymethyl cellulose (羧甲基纤维素) 示例: 以口服配方 1 (悬浮于 0.5% CMC Na)为例说明, 如果要配制 100 mL 2.5 mg/mL 的工作液, 您可以先取0.5g CMC Na并将其溶解于100mL ddH2O中,得到0.5%CMC-Na澄清溶液;然后将250 mg待测化合物加到100 mL前述 0.5%CMC Na溶液中,得到悬浮液。 View More
口服配方 3: 溶解于 PEG400 (聚乙二醇400) 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.5047 mL | 12.5235 mL | 25.0470 mL | |
| 5 mM | 0.5009 mL | 2.5047 mL | 5.0094 mL | |
| 10 mM | 0.2505 mL | 1.2523 mL | 2.5047 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1





























 463611831
463611831
