| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 1mg |
|
||
| Other Sizes |
|

| 靶点 |
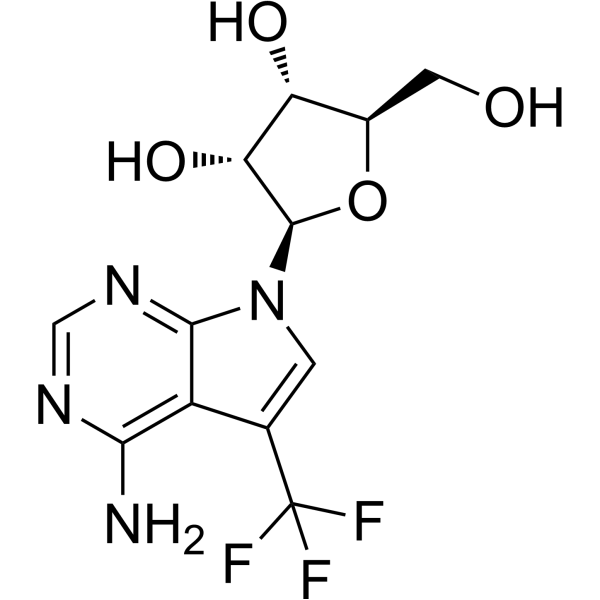
MTr1 (2'-O-ribose methyltransferase 1)
|
|---|---|
| 体外研究 (In Vitro) |
在第三步中,研究人员评估了哪些已鉴定的化合物在最低浓度下具有抑制作用,并最终确定了最有效的化合物是Trifluoromethyl-tubercidin/三氟甲基tubercidin(TFMT)(图2A在图像查看器中打开,图S8、A和B)。我们分别通过计算机对接和体外热位移分析证实了TFMT与MTr1和重组MTr1的SAM结合袋的结合(图2BOpen in image viewer和图S8C)。此外,我们证实TFMT抑制重组MTr1蛋白的MTase活性(图S8D)。我们还证实,TFMT对IAV和IBV有效,但对HAZV或STBV无效(图2COpen在图像查看器中),与图1Open在图像浏览器和图S6中显示的MTr1缺乏症表型完全匹配。TFMT对IAV感染的中位抑制浓度(IC50)为0.30μM,在有效浓度范围内没有观察到明显的体外毒性,如水溶性四唑(WST)-8细胞存活率测定所示(图2D在图像查看器中打开)。TFMT治疗在感染后3至4小时给药时也大大抑制了IAV的复制;然而,如果稍后给药,效果会减弱或不可见(图S8E)[1]。
Trifluoromethyl-tubercidin/三氟甲基结核菌素/TFMT[1]的抗流感疗效 接下来,研究人员检测了正常人支气管上皮(NHBE)细胞中三氟甲基结核菌素/TFMT的抗IAV活性。TFMT治疗以剂量依赖的方式显著降低了IAV(H1N1,PR8)的RNA和蛋白质水平。组织学分析还显示,TFMT处理的NHBE细胞中IAV NP水平显著降低,且无细胞毒性。TFMT处理没有抑制HAZV复制,表明即使在人类原代细胞中,该化合物对某些病毒的特异性疗效也得以保留。由于该化合物在人NHBE细胞中有效,这促使我们将TFMT作为离体环境在人肺外植体中进行评估(图S9A)。我们用IAV(H1N1,2019年的季节性分离株)感染肺组织,并在感染后1、24、48和72小时用空斑试验测定上清液中的病毒滴度。如图3A所示,在图像查看器中打开时,感染后48或72小时,未处理样本的滴度增加了>105个斑块形成单位(PFU)/ml,而TFMT处理的肺外植体的滴度保持在<103个PFU/ml,表明治疗抑制了100到1000倍。所有六个独立供体的滴度之和显示了对照组和TFMT治疗之间的差异——培养上清液中的IAV滴度降低。感染后12小时IAV的TFMT治疗显著损害了人肺外植体中的病毒生长(图S9C)。与观察到的病毒滴度一致,在用TFMT治疗的IAV感染的肺组织中没有观察到IAV NP阳性细胞或形态学变化。这些结果表明,TFMT抑制季节性IAV分离物的体外复制,并显示出临床转化的潜力。 TFMT/三氟甲基结核菌素/Trifluoromethyl-tubercidin抑制IAV帽夺[1] 如图4A所示,在图像查看器中打开,TFMT处理对IAV复制的影响与IFIT1依赖的RNA或RIG-I或MDA5信号隔离无关。我们发现RIG-I-MTr1双KO细胞或IFIT1-MTr1双KO细胞中没有IAV的复制,与MTr1 KO细胞相似(图S11、A和B)。此外,JAK抑制剂托法替尼阻断IFN信号传导时,MTr1 KO细胞中的IAV复制并不伴随(图S12、C和D)。已知缺失非结构蛋白1(IAVΔNS1)的甲型流感病毒可诱导高IFN反应,在没有IFN和ISG(干扰素刺激基因)诱导的情况下,其在MTr1缺陷细胞中的复制被阻止,但被MTr1过表达挽救(图S2、D和E)。TFMT治疗在A549细胞和PBMC中均未诱导IFN-β或抗病毒ISG。这些结果证实,观察到的抗病毒作用不依赖于抗病毒IFN反应的激活。TFMT治疗不会改变IFN敏感的非帽状RNA病毒的复制,如仙台病毒(SeV)、水疱性口炎病毒(VSV)和脑心肌炎病毒(EMCV),同样,在MTr1 KO细胞中(图S13)也是如此。TFMT处理会损害特异性从U2剪接体snRNA中捕获的IAV mRNA(片段1)的表达,与MTr1缺乏的影响相似。因此,我们得出结论,TFMT治疗通过直接影响IAV的夺帽活性而不是通过免疫调节来抑制IAV的复制。 |
| 体内研究 (In Vivo) |
研究人员在小鼠身上测试了这种化合物的体内功效。他们首先证实,TFMT/三氟甲基结核菌素在IAV感染的小鼠细胞系LA-4中显示出抑制活性,尽管其效力(IC50=7.7μM)低于人类细胞(图S10A)。接下来,我们评估了小鼠每天一次鼻内接种2天的体内毒性。尽管用亲本化合物tubercidin治疗导致小鼠体重大幅减轻,但我们没有观察到所选衍生物TFMT在肺部有任何体重减轻或细胞毒性(图3D在图像查看器中打开,图S10B)。最后,我们在感染IAV后2天检查了TFMT治疗的效果。此时,TFMT治疗显著降低了小鼠肺部NP和PB2 mRNA水平,表明三氟甲基取代的tubercidin将体内毒性降低到我们无法检测到的水平,但保留了抗IAV的功效(图3E在图像查看器中打开)。我们还证实了在这种情况下,巴洛西韦(BXM)在体内的抗病毒功效(图S10C)。综上所述,TFMT显示出在所有测试系统中抑制IAV复制的潜力,包括体外的人细胞系和NHBE细胞、体外的人肺外植体和体内的小鼠[1]。
|
| 参考文献 | |
| 其他信息 |
正粘病毒和布尼亚病毒会窃取宿主RNA的5'端帽状结构来启动自身的转录,这一过程被称为“帽状结构窃取”。我们发现,宿主2'-O-核糖甲基转移酶1 (MTr1) 对帽状结构的RNA修饰对于甲型和乙型流感病毒复制的启动至关重要,但对其他帽状结构窃取病毒则并非如此。我们通过计算机化合物筛选和功能分析,鉴定出一种链霉菌天然产物衍生物——三氟甲基结核菌素 (TFMT),它通过与MTr1的S-腺苷-L-蛋氨酸结合口袋相互作用来抑制MTr1,从而限制流感病毒的复制。机制上,TFMT会削弱宿主帽状RNA与病毒聚合酶碱性蛋白2亚基的结合,这一现象在人肺组织外植体和体内小鼠中均有发现。 TFMT 与已获批准的抗流感药物具有协同作用。[1]
目前已有针对流感病毒蛋白的获批药物;然而,所有药物均已出现耐药病毒突变株。宿主靶向抗病毒药物诱导耐药性的可能性较低,宿主丝裂原活化蛋白激酶抑制剂 ATR-002 已被证明对包括流感病毒和 SARS-CoV-2 在内的多种 RNA 病毒具有广泛的疗效,其作用机制是通过抑制病毒复制和调节炎症反应。在本研究中,我们展示了细胞 RNA 甲基转移酶抑制剂 三氟甲基结核菌素/TFMT 的抗流感疗效。考虑到长期靶向宿主因子可能产生的毒性,降低剂量并与其他已获批准的抗病毒药物(如BXM和奥司他韦)联合使用是切实可行的。 我们发现TFMT是一种高度特异性且无毒的MTr1抑制剂,能够特异性地抑制依赖于帽状结构捕获的病毒(如IAV和IBV)的复制(图S19)。总的来说,我们的数据表明,TFMT对MTr1帽状结构捕获的抑制作用能够特异性地抑制多种IAV和IBV毒株的复制,包括季节性H1N1分离株和一种高致病性且对BXM耐药的禽流感病毒。从机制上讲,TFMT导致MTr1功能障碍和cap0 RNA积累,从而损害病毒聚合酶亚基PB2与宿主帽状RNA的结合,进而阻碍IAV聚合酶介导的帽状结构捕获和病毒RNA合成。 TFMT 与 BXM 具有协同作用,因为这两种药物分别靶向不同的聚合酶亚基 PB2 和 PA。TFMT 依赖的 IAV 抑制作用不依赖于 RIG-I 和 IFIT1 介导的先天免疫反应,并且 TFMT 对干扰素敏感病毒(如 VSV 和 EMCV)的复制没有影响。流感病毒和 THOV 的 PB2 亚基比较显示,IAV PB2 的帽 RNA 结合区(例如 N1-2′-O-Me 相互作用氨基酸)的一级结构在 IBV PB2 中保守,但在 ICV、IDV 或 THOV PB2 中不保守(图 S14B)。此外,IAV 需要 10 至 13 个核苷酸才能捕获帽结构,而据报道 THOV 捕获的是 5′ 末端的 m7G 帽残基。病毒聚合酶在夺帽作用上的机制差异可能解释了 TFMT 限制对 IAV 和 IBV 的特异性。[1] |
| 分子式 |
C12H13F3N4O4
|
|---|---|
| 分子量 |
334.251232862473
|
| 精确质量 |
334.088
|
| CAS号 |
1854086-05-7
|
| PubChem CID |
118636125
|
| 外观&性状 |
White to off-white solid powder
|
| LogP |
-0.5
|
| tPSA |
127
|
| 氢键供体(HBD)数目 |
4
|
| 氢键受体(HBA)数目 |
10
|
| 可旋转键数目(RBC) |
2
|
| 重原子数目 |
23
|
| 分子复杂度/Complexity |
443
|
| 定义原子立体中心数目 |
4
|
| SMILES |
C1=C(C2=C(N=CN=C2N1[C@H]3[C@@H]([C@@H]([C@H](O3)CO)O)O)N)C(F)(F)F
|
| InChi Key |
RSOXZOFDCJMRMK-IOSLPCCCSA-N
|
| InChi Code |
InChI=1S/C12H13F3N4O4/c13-12(14,15)4-1-19(10-6(4)9(16)17-3-18-10)11-8(22)7(21)5(2-20)23-11/h1,3,5,7-8,11,20-22H,2H2,(H2,16,17,18)/t5-,7-,8-,11-/m1/s1
|
| 化学名 |
(2R,3R,4S,5R)-2-[4-amino-5-(trifluoromethyl)pyrrolo[2,3-d]pyrimidin-7-yl]-5-(hydroxymethyl)oxolane-3,4-diol
|
| 别名 |
Trifluoromethyl-tubercidin; 1854086-05-7; (2R,3R,4S,5R)-2-[4-amino-5-(trifluoromethyl)pyrrolo[2,3-d]pyrimidin-7-yl]-5-(hydroxymethyl)oxolane-3,4-diol; TFMT?; SCHEMBL17406905; RSOXZOFDCJMRMK-IOSLPCCCSA-N; NSC793694; NSC-793694;
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
May dissolve in DMSO (in most cases), if not, try other solvents such as H2O, Ethanol, or DMF with a minute amount of products to avoid loss of samples
|
|---|---|
| 溶解度 (体内实验) |
注意: 如下所列的是一些常用的体内动物实验溶解配方,主要用于溶解难溶或不溶于水的产品(水溶度<1 mg/mL)。 建议您先取少量样品进行尝试,如该配方可行,再根据实验需求增加样品量。
注射用配方
注射用配方1: DMSO : Tween 80: Saline = 10 : 5 : 85 (如: 100 μL DMSO → 50 μL Tween 80 → 850 μL Saline)(IP/IV/IM/SC等) *生理盐水/Saline的制备:将0.9g氯化钠/NaCl溶解在100 mL ddH ₂ O中,得到澄清溶液。 注射用配方 2: DMSO : PEG300 :Tween 80 : Saline = 10 : 40 : 5 : 45 (如: 100 μL DMSO → 400 μL PEG300 → 50 μL Tween 80 → 450 μL Saline) 注射用配方 3: DMSO : Corn oil = 10 : 90 (如: 100 μL DMSO → 900 μL Corn oil) 示例: 以注射用配方 3 (DMSO : Corn oil = 10 : 90) 为例说明, 如果要配制 1 mL 2.5 mg/mL的工作液, 您可以取 100 μL 25 mg/mL 澄清的 DMSO 储备液,加到 900 μL Corn oil/玉米油中, 混合均匀。 View More
注射用配方 4: DMSO : 20% SBE-β-CD in Saline = 10 : 90 [如:100 μL DMSO → 900 μL (20% SBE-β-CD in Saline)] 口服配方
口服配方 1: 悬浮于0.5% CMC Na (羧甲基纤维素钠) 口服配方 2: 悬浮于0.5% Carboxymethyl cellulose (羧甲基纤维素) 示例: 以口服配方 1 (悬浮于 0.5% CMC Na)为例说明, 如果要配制 100 mL 2.5 mg/mL 的工作液, 您可以先取0.5g CMC Na并将其溶解于100mL ddH2O中,得到0.5%CMC-Na澄清溶液;然后将250 mg待测化合物加到100 mL前述 0.5%CMC Na溶液中,得到悬浮液。 View More
口服配方 3: 溶解于 PEG400 (聚乙二醇400) 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.9918 mL | 14.9589 mL | 29.9177 mL | |
| 5 mM | 0.5984 mL | 2.9918 mL | 5.9835 mL | |
| 10 mM | 0.2992 mL | 1.4959 mL | 2.9918 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
 Fusion Inhibitory Peptide (ZD-Phe-Phe-Gly-OH; FIP; Virus Replication Inhibitory Peptide)
Fusion Inhibitory Peptide (ZD-Phe-Phe-Gly-OH; FIP; Virus Replication Inhibitory Peptide)
 ZIKV-IN-8
ZIKV-IN-8
 PAN endonuclease-IN-1
PAN endonuclease-IN-1
 Urolithin M5 (Decarboxyellagic acid)
Urolithin M5 (Decarboxyellagic acid)
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 463611831
463611831
