| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 100mg |
|
||
| 500mg |
|
||
| 1g |
|
||
| Other Sizes |
|

| 靶点 |
Methyl jasmonate targets plant signaling pathways involved in defense responses and secondary metabolite biosynthesis. As a plant hormone, it regulates the expression of genes involved in plant defense, stress responses, and development. In biomedical research, methyl jasmonate has been studied for its anticancer and anti-inflammatory effects. Its mechanism of action in mammalian cells involves modulation of cell signaling pathways such as MAPK and NF-κB, though its precise molecular targets have not been definitively identified. Its plant hormone activity and potential biomedical applications make it a valuable tool for plant biology and pharmacology research.
|
|---|---|
| 体外研究 (In Vitro) |
体外实验表明,茉莉酸甲酯在哺乳动物细胞系中具有抗癌和抗炎活性。它能诱导癌细胞凋亡并抑制其增殖。该化合物的活性呈浓度依赖性,有效浓度通常在微摩尔范围内。在植物细胞培养中,茉莉酸甲酯能诱导次生代谢产物的产生并激活防御相关基因。其在植物和哺乳动物系统中的双重活性使其成为研究植物生物学和药理学的宝贵工具。特定癌细胞系的详细IC50值可在已发表的文献中找到。
|
| 体内研究 (In Vivo) |
在体内,茉莉酸甲酯已在癌症和炎症的临床前模型中进行了研究。其诱导细胞凋亡和抑制肿瘤生长的能力使其成为一种有前景的抗癌药物候选物。然而,公开资料中关于其体内疗效和药代动力学特征的详细数据有限。该化合物目前主要用作植物诱导和生物医学研究的工具。需要开展进一步的研究,以全面阐明其体内治疗潜力、给药方案和安全性。
|
| 酶活实验 |
茉莉酸甲酯的体外抗癌活性测定通常使用癌细胞系。将细胞接种于96孔板中,并用不同浓度的化合物(通常为1至100 µM)处理24至72小时。采用MTT或CellTiter-Glo法评估细胞活力。通过Annexin V/PI染色和caspase活性测定定量细胞凋亡。采用碘化丙啶染色和流式细胞术分析细胞周期分布。在植物研究中,用茉莉酸甲酯处理植物细胞培养物或组织,并通过HPLC或LC-MS测定次生代谢产物的生成。采用qRT-PCR评估基因表达。每次实验均设置阳性对照和阴性对照。
|
| 细胞实验 |
在体外细胞实验中,将癌细胞系或植物细胞培养物用浓度为1至100 µM的茉莉酸甲酯处理24至72小时。采用MTT或CellTiter-Glo法评估细胞活力。通过Annexin V/PI染色和caspase活性测定定量细胞凋亡。采用ELISA法检测炎症标志物(TNF-α、IL-6)。在植物研究中,采用HPLC或LC-MS法测定次生代谢产物的生成。采用qRT-PCR法评估基因表达。所有实验均设置适当的对照(溶剂对照、未处理细胞),并重复三次。
|
| 动物实验 |
在体内疗效研究中,通常使用啮齿动物癌症或炎症模型。茉莉酸甲酯以1~50 mg/kg的剂量口服或腹腔注射给药,通常每日一次或两次。监测肿瘤生长情况,并检测血液和组织中的炎症标志物。在植物研究中,将茉莉酸甲酯施用于植物,并评估其防御反应和次生代谢产物的产生。所有动物实验均应按照机构指南进行。
|
| 药代性质 (ADME/PK) |
茉莉酸甲酯的药代动力学性质已得到部分表征。该化合物的分子量为224.30,LogP值为2.5。口服或腹腔注射后,其吸收中等,达峰时间(Tmax)为1-3小时。血浆半衰期估计为2-4小时。该化合物分布于包括肝脏和肾脏在内的组织中。代谢主要在肝脏进行,氧化和结合是主要代谢途径。该化合物主要通过胆汁和肾脏排泄。需要进一步的药代动力学研究以进行全面表征。
|
| 毒性/毒理 (Toxicokinetics/TK) |
相互作用
研究人员研究了茉莉酸甲酯和蔗糖联合作用对葡萄细胞悬浮液中防御相关基因表达、芪类化合物和花青素生成的影响。茉莉酸甲酯/蔗糖处理有效刺激了苯丙氨酸解氨酶、查尔酮合酶、芪类合酶、UDP-葡萄糖:黄酮-O-葡萄糖基转移酶、蛋白酶抑制剂和几丁质酶等基因的表达,并诱导了细胞内云杉化合物和花青素的积累,以及细胞外培养基中反式白藜芦醇和云杉化合物的积累…… 辣椒悬浮细胞培养物被用于评估环糊精和茉莉酸甲酯作为防御反应诱导剂的效果。诱导的防御反应包括倍半萜烯和植物甾醇的积累,以及病程相关蛋白的激活,从而在诱导过程中增强和改变细胞壁结构,并保护细胞免受生物胁迫。结果表明,环糊精和茉莉酸甲酯的共同添加诱导了两种倍半萜烯——芳香树脂和索拉维酮的生物合成。这种反应表现出显著的协同效应,因为当两种诱导剂同时存在时,这些化合物的含量增加远高于单独使用时。由于累加效应,联合处理也诱导了植物甾醇的生物合成。同样,外源施用茉莉酸甲酯也诱导了疾病相关蛋白的积累。细胞外蛋白质组学分析揭示了与PR1和PR4同源的氨基酸序列、NtPRp27样蛋白、I类几丁质酶、过氧化物酶、水解酶LEXYL1和LEXYL2、阿拉伯糖苷酶、果胶酶、油桃素IV以及富含亮氨酸重复序列蛋白的存在,表明茉莉酸甲酯介导了辣椒(C. annuum)中防御相关基因产物的表达。除这些茉莉酸甲酯诱导的蛋白外,在辣椒的对照和诱导细胞培养物中均发现了其他PR蛋白。这些酶,包括IV类几丁质酶、β-1,3-葡聚糖酶、甜蛋白样酶和过氧化物酶,表明它们的表达主要为组成型表达,因为它们参与植物的生长、发育和防御过程。硼是植物必需的微量元素,但土壤中过量的硼对某些植物(例如青蒿)具有植物毒性,青蒿的地上部分含有青蒿素(一种重要的抗疟疾药物)。青蒿素是一种倍半萜内酯,其分子内含有过氧化桥……本研究旨在确定外源施用茉莉酸甲酯(MeJA)是否能够抵消土壤中过量硼胁迫(B)的不利影响。结果表明,硼毒性诱导了氧化胁迫,并显著降低了茎高、鲜重和干重。土壤中过量的硼降低了叶片的净光合速率、气孔导度、胞间二氧化碳浓度和总叶绿素含量。相反,叶面喷施茉莉酸甲酯(MeJA)改善了受胁迫和未受胁迫植物的生长和光合效率。过量的硼还会增加抗氧化酶(如过氧化氢酶、过氧化物酶和超氧化物歧化酶)的活性……对受胁迫的植物施用茉莉酸甲酯(MeJA)可以减少脂质过氧化,刺激抗氧化酶的合成,并提高青蒿素的含量和产量。因此,可以得出结论,MeJA 可用于缓解硼中毒,并提高青蒿中青蒿素的含量和产量。 茉莉酸甲酯的临床前毒理学研究有限。在啮齿动物的急性毒性研究中,该化合物在高达 50 mg/kg 的剂量下耐受性良好,未观察到明显的不良反应。在重复给药研究中,尚未明确确定未观察到不良反应剂量(NOAEL)。在药理剂量下,未报告明显的器官毒性或血液学异常。该化合物在标准体外试验中未显示遗传毒性。该化合物的安全性数据支持进一步的临床前开发,但仍需进行全面的毒理学研究,以充分评估其安全性,从而推进临床应用。该化合物仅供研究使用,尚未获准用于人体。 |
| 参考文献 | |
| 其他信息 |
茉莉酸甲酯(-)是茉莉酸的甲酯衍生物,属于茉莉酸酯类,是一种植物代谢物和植物激素。它是一种茉莉酸酯,也是一种甲酯,是茉莉酸酯衍生物家族的成员。据报道,茉莉酸甲酯存在于马铃薯(Solanum tuberosum)、雷公藤(Tripterygium wilfordii)和其他具有相关数据的生物体中。作用机制:作者以链格孢菌(Alternaria alternata)的致病型及其AAL毒素/番茄互作系统为模型系统,论证了茉莉酸(JA)在植物对利用宿主特异性毒素作为毒力效应因子的病原体的易感性中的潜在作用。与野生型(WT)品种相比,JA生物合成缺陷的def1突变体在Aa致病番茄植株中表现出疾病发展和植物生长受到抑制。外源施用茉莉酸甲酯 (MeJA) 可恢复 def1 突变体中的病原症状,并加剧野生型植株的病情。另一方面,AAL 毒素在 def1 和野生型植株中均诱导了类似的坏死性细胞死亡,而施用 MeJA 并不影响毒素诱导的细胞死亡程度。这些结果表明,JA 依赖性信号通路不参与番茄对 Aa 病原体的基础防御反应,但可能以不依赖于毒素的方式影响病原体的耐受性。进一步的数据表明,茉莉酸 (JA) 可促进产毒病原体和坏死性病原体对番茄的侵染,并且病原体可能利用 JA 信号通路成功侵染。……WRKY 是一种植物特异性转录因子,也是其非寄主拟南芥中的鞭毛蛋白诱导基因之一。接种含有AvrRpt2的不亲和病原菌丁香假单胞菌DC3000 (Pto)和非寄主病原菌可诱导WRKY41表达……过表达WRKY41的拟南芥对野生型Pto表现出增强的抗性,但对胡萝卜素欧文氏菌EC1的易感性增加。过表达WRKY41的拟南芥组成型表达PR5基因,但抑制了茉莉酸甲酯诱导的PDF1.2基因表达。这些结果表明,WRKY41可能是水杨酸和茉莉酸信号通路相互作用的关键调控因子。
诱导细胞死亡是植物抵御病原体的重要组成部分。大量研究报道了植物激素在病原体诱导的细胞死亡中的作用,但茉莉酸(JA)尚未被鉴定为该反应的调控因子。本文报道了本氏烟草同源框1 (NbHB1) 在茉莉酸 (JA) 信号通路中病原体诱导的细胞死亡中的功能。研究人员分别利用农杆菌介导的瞬时过表达和病毒诱导的基因沉默技术,通过功能获得和功能丧失实验分析了NbHB1在细胞死亡中的作用。采用逆转录聚合酶链式反应 (RT-PCR) 检测了病原体接种和不同处理后NbHB1的表达水平。结果表明,强毒和弱毒细菌病原体的感染均能上调NbHB1的转录水平。NbHB1的异位表达加速了黑暗、茉莉酸甲酯或病原体接种后的细胞死亡。相反,当NbHB1被沉默时,病原体诱导的细胞死亡被延迟。NbCOI1的沉默也延迟了NbHB1诱导的细胞死亡,表明JA介导的信号通路至关重要。 NbHB1结构域缺陷蛋白的过表达表明,同源结构域、亮氨酸拉链结构域和部分可变的N端区域对NbHB1的功能至关重要。这些结果强烈提示NbHB1通过茉莉酸(JA)介导的信号通路在病原体诱导的植物细胞死亡中发挥作用。本研究利用高通量Illumina测序技术鉴定红豆杉(Taxus chinensis)细胞中的miRNA,以研究紫杉烷类诱导剂茉莉酸甲酯(MJ)对miRNA表达的影响。在包含约660万条序列的数据集中,鉴定出58个miRNA,分属25个家族。其中大多数在被子植物和裸子植物中均保守。然而,有两个miRNA(miR1310和miR1314)似乎是裸子植物特有的,其中miR1314可能以簇状形式存在。MJ处理显著影响了特定miRNA的表达;来自7个不同家族的14个miRNA(miR156、miR168、miR169、miR172、miR396、miR480和miR1310)表达下调,而来自2个家族的3个miRNA(miR164和miR390)表达上调。有关茉莉酸甲酯作用机制(共13种)的更完整数据,请访问HSDB记录页面。茉莉酸甲酯(MeJA)是茉莉酸的甲酯,茉莉酸是一种植物激素,可调节植物的防御反应和次生代谢产物的生物合成。它在生物医学研究中具有抗癌和抗炎活性。该化合物尚未获准用于人体,也未进入临床试验。它仅作为高纯度研究试剂供实验室使用。其植物激素活性和生物医学潜力使其成为植物生物学和药理学研究的重要工具。 |
| 分子式 |
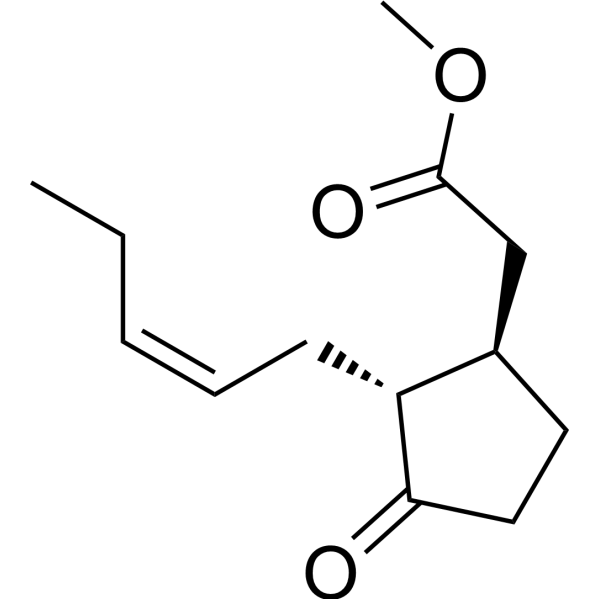
C13H20O3
|
|---|---|
| 分子量 |
224.2961
|
| 精确质量 |
224.141
|
| CAS号 |
1211-29-6
|
| PubChem CID |
5281929
|
| 外观&性状 |
Colorless liquid
|
| 密度 |
1.0±0.1 g/cm3
|
| 沸点 |
302.9±15.0 °C at 760 mmHg
|
| 熔点 |
25 °C
|
| 闪点 |
128.6±20.4 °C
|
| 蒸汽压 |
0.0±0.6 mmHg at 25°C
|
| 折射率 |
1.469
|
| LogP |
2.12
|
| tPSA |
43.37
|
| 氢键供体(HBD)数目 |
0
|
| 氢键受体(HBA)数目 |
3
|
| 可旋转键数目(RBC) |
6
|
| 重原子数目 |
16
|
| 分子复杂度/Complexity |
281
|
| 定义原子立体中心数目 |
2
|
| SMILES |
CC/C=C\C[C@@H]1[C@H](CCC1=O)CC(=O)OC
|
| InChi Key |
GEWDNTWNSAZUDX-WQMVXFAESA-N
|
| InChi Code |
InChI=1S/C13H20O3/c1-3-4-5-6-11-10(7-8-12(11)14)9-13(15)16-2/h4-5,10-11H,3,6-9H2,1-2H3/b5-4-/t10-,11-/m1/s1
|
| 化学名 |
methyl 2-[(1R,2R)-3-oxo-2-[(Z)-pent-2-enyl]cyclopentyl]acetate
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
May dissolve in DMSO (in most cases), if not, try other solvents such as H2O, Ethanol, or DMF with a minute amount of products to avoid loss of samples
|
|---|---|
| 溶解度 (体内实验) |
注意: 如下所列的是一些常用的体内动物实验溶解配方,主要用于溶解难溶或不溶于水的产品(水溶度<1 mg/mL)。 建议您先取少量样品进行尝试,如该配方可行,再根据实验需求增加样品量。
注射用配方
注射用配方1: DMSO : Tween 80: Saline = 10 : 5 : 85 (如: 100 μL DMSO → 50 μL Tween 80 → 850 μL Saline)(IP/IV/IM/SC等) *生理盐水/Saline的制备:将0.9g氯化钠/NaCl溶解在100 mL ddH ₂ O中,得到澄清溶液。 注射用配方 2: DMSO : PEG300 :Tween 80 : Saline = 10 : 40 : 5 : 45 (如: 100 μL DMSO → 400 μL PEG300 → 50 μL Tween 80 → 450 μL Saline) 注射用配方 3: DMSO : Corn oil = 10 : 90 (如: 100 μL DMSO → 900 μL Corn oil) 示例: 以注射用配方 3 (DMSO : Corn oil = 10 : 90) 为例说明, 如果要配制 1 mL 2.5 mg/mL的工作液, 您可以取 100 μL 25 mg/mL 澄清的 DMSO 储备液,加到 900 μL Corn oil/玉米油中, 混合均匀。 View More
注射用配方 4: DMSO : 20% SBE-β-CD in Saline = 10 : 90 [如:100 μL DMSO → 900 μL (20% SBE-β-CD in Saline)] 口服配方
口服配方 1: 悬浮于0.5% CMC Na (羧甲基纤维素钠) 口服配方 2: 悬浮于0.5% Carboxymethyl cellulose (羧甲基纤维素) 示例: 以口服配方 1 (悬浮于 0.5% CMC Na)为例说明, 如果要配制 100 mL 2.5 mg/mL 的工作液, 您可以先取0.5g CMC Na并将其溶解于100mL ddH2O中,得到0.5%CMC-Na澄清溶液;然后将250 mg待测化合物加到100 mL前述 0.5%CMC Na溶液中,得到悬浮液。 View More
口服配方 3: 溶解于 PEG400 (聚乙二醇400) 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 4.4583 mL | 22.2916 mL | 44.5831 mL | |
| 5 mM | 0.8917 mL | 4.4583 mL | 8.9166 mL | |
| 10 mM | 0.4458 mL | 2.2292 mL | 4.4583 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
 AOH-1996
AOH-1996
 hMAO-B-IN-3
hMAO-B-IN-3
 Flupyrimin
Flupyrimin
 阿曲库铵
阿曲库铵
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 463611831
463611831
