| 规格 | 价格 | |
|---|---|---|
| 500mg | ||
| 1g | ||
| Other Sizes |

| 毒性/毒理 (Toxicokinetics/TK) |
毒性概述
鉴定与用途:箭毒蛙毒素是一类神经毒性甾体生物碱,最初从哥伦比亚箭毒蛙中分离得到,后来在新几内亚的某些雀形目鸟类中也发现了该毒素。目前认为这两种脊椎动物都不会从头合成该毒素,而是可能从食物中获取。人体研究:箭毒蛙毒素是电压门控钠通道的强效调节剂,会导致神经和肌肉不可逆去极化、心房颤动、心律失常,最终导致心力衰竭。自发现以来,野外研究人员也报告称,皮肤接触该毒素后会出现麻木感。动物实验:当P. terribilis被圈养时,其体内的箭毒蛙毒素水平往往会降低,但即使被圈养长达6年,这些蛙的毒性仍然至少是当地居民用于制作毒箭的其他叶蛙属(Phyllobates)物种的五倍。在圈养至成熟的F1代蛙中未检测到箭毒蛙毒素。从野生蛙和无毒的F1代蛙中提取的神经和肌肉组织均对箭毒蛙毒素不敏感。控制钠通道激活和通透性的调控位点似乎仅发生了轻微改变,以防止与箭毒蛙毒素相互作用,但仍然对其他钠离子通道激活剂(藜芦碱、灰毒素)敏感,而蛙类在自然界中并未接触到这些激活剂。 相互作用 此前已发现两种不同类型的局部麻醉剂(LA)能够阻断箭毒蛙毒素(BTX)修饰的钠离子通道:I型局部麻醉剂,例如可卡因和布比卡因,优先与开放通道相互作用;而II型局部麻醉剂,例如苯佐卡因和三卡因,则与失活通道相互作用。本文中,我们描述了我们对第三种局部麻醉剂——丁卡因的研究。丁卡因是一种双重阻滞剂,它能与关闭的通道强力结合,但在膜去极化时也能与开放的通道有一定程度的结合。通过稳态失活测定和剂量反应曲线确定了丁卡因对BTX修饰的Na+通道的增强失活作用。在-70 mV时,稳态失活达到最大值,此时的半数抑制浓度(IC50)估计为5.2 μM,Hill系数为0.98,表明一个丁卡因分子与一个失活的通道结合。当膜去极化时,丁卡因也能有效地与Na+通道相互作用;在+50 mV时,IC50估计为39.5 μM,Hill系数为0.94。出乎意料的是,带电荷的丁卡因是阻断失活通道的主要活性形式。此外,胞外Na+离子似乎能拮抗丁卡因对失活通道的阻断作用。与这些结果一致,N-丁基丁卡因季铵盐(一种永久带电荷的丁卡因衍生物)仍然是一种强效的失活增强剂。另一种丁卡因衍生物,苯环上缺少4-丁氨基官能团的2-(二甲基氨基)乙基苯甲酸酯,其阻断作用比丁卡因弱约100倍。我们推测:1)失活增强剂的结合位点位于Na+渗透通路内;2)外部Na+离子通过静电排斥拮抗失活增强剂的阻断作用;3)苯环上的4-丁基氨基官能团对于阻断和增强失活至关重要;4)Na+孔内可能存在失活增强剂和开放通道阻滞剂的重叠结合位点。 目的:为了研究不同钠通道亚型对全身麻醉剂的反应,我们检测了戊巴比妥(一种硫喷妥钠的类似物)对人骨骼肌单个钠通道的影响,并将其与来自人脑和人心室肌通道的现有数据进行了比较。方法:将人骨骼肌钠通道制备物整合到平面脂双层中,并在钠通道激活剂河豚毒素存在下,检测单个钠通道的稳态行为及其对戊巴比妥的反应。在加入戊巴比妥(0.34-1.34 mM)前后记录单通道电流。结果:在对称的500 mM NaCl溶液中,人骨骼肌钠通道的平均单通道电导为21.0 ± 0.6 pS,通道开放分数为0.96 ± 0.04。激活中点电位为-96.2 ± 1.6 mV。胞外河豚毒素在0 mV时以60 nM的半数最大浓度(k1/2)阻断该通道。戊巴比妥以浓度依赖的方式降低了单个骨骼肌钠通道的时间平均电导(半数抑制浓度[IC50] = 0.66 mM)。稳态激活电位向更负的电位移动(0.67 mM 戊巴比妥时为 -16.7 mV)。结论:在平面脂双层系统中,骨骼肌钠通道的某些电生理特性与心肌或中枢神经组织中的钠通道显著不同。与对照数据相反,这些不同的人类钠通道亚型对全身麻醉剂戊巴比妥表现出相同的定性和定量反应。这些效应对整体麻醉效果的影响取决于各个通道在其神经元网络中的作用,但抑制中枢神经系统和外周钠通道可能会增强全身麻醉效果。 我们研究了普鲁卡因胺对牛心脏和鼠骨骼肌中箭毒杆菌毒素 (BTX) 激活的钠通道的作用。当应用于细胞内侧时,普鲁卡因胺可诱导快速的开放通道阻滞。我们使用振幅分布分析估算了速率常数。膜去极化增加了阻滞速率并减缓了阻滞解除。心肌和骨骼肌通道的速率常数在幅度和电压依赖性方面均相似。从定性上看,这种阻滞类似于利多卡因引起的快速开放通道阻滞,但普鲁卡因胺的效力大约低七倍。分子建模表明,普鲁卡因胺和利多卡因效力差异可能源于它们芳香环的相对取向,或芳胺键结构的差异。对于心脏通道,普鲁卡因胺降低了通道向长寿命关闭状态转变的频率,该状态表现出失活的特征。动力学鉴定的关闭状态的平均持续时间未受影响。快速阻滞和慢速关闭抑制的程度呈正相关。内源性应用利多卡因衍生物QX-314(也是一种快速阻滞剂)产生了类似的效果。因此,药物与快速阻滞位点的结合似乎抑制了BTX激活的心脏通道的失活。 本研究旨在探讨箭毒蛙毒素(BTX)对心肌细胞中Na+通道的修饰特征,包括通道门控和动力学的变化以及对局部麻醉剂阻滞的敏感性。我们采用膜片钳技术的全细胞模式测量豚鼠心肌细胞中的Na+电流。为了保持良好的电压控制,我们降低了细胞外Na+浓度和温度(5-10 mM,17℃)。我们的研究结果表明:1)BTX可改变心脏钠离子通道的稳态(非失活)成分,导致钠离子通道中出现显著的稳态(非失活)成分;2)BTX对心脏钠离子通道的修饰使激活电位向更负的方向移动,并降低最大钠离子通道阻滞阈值(gNa)和对钠离子的选择性;3)BTX与其在心脏钠离子通道上的受体结合,降低了局部麻醉药对其结合位点的亲和力;4)BTX修饰的通道表现出局部麻醉药的使用依赖性阻滞。局部麻醉药对BTX修饰的钠离子通道阻滞效力降低可能是由于BTX和局部麻醉药在其钠离子通道结合位点上的变构相互作用所致。我们观察到局部麻醉药的使用依赖性阻滞在BTX修饰的钠离子通道中仍然存在,这表明这种额外的阻滞形式可以在几乎不存在失活状态的情况下发生。因此,在这些条件下,使用依赖性阻滞的发生似乎主要依赖于局部麻醉剂与激活的Na+通道的结合。 有关巴曲毒素(共6种)的更多相互作用(完整)数据,请访问HSDB记录页面。 非人类毒性值 小鼠静脉注射LD50:0.002 mg/kg 小鼠皮肤给药LD50:2 μg/kg |
|---|---|
| 参考文献 | |
| 其他信息 |
已有报道称,金带蛙(Phyllobates aurotaenia)体内存在蛙毒素(Batrachotoxin),并有相关数据可供参考。
蛙毒素是一种强效的钠通道激活剂,已在蛙皮提取物中被发现。 作用机制 蛙毒素是电压门控钠通道的强效调节剂,可导致神经和肌肉的不可逆去极化、颤动、心律失常,最终导致心力衰竭。自发现以来,野外研究人员也报告称,皮肤接触该毒素后会出现麻木感。出于对这一现象的好奇,我们研究了蛙毒素对电压门控钠通道Nav1.8的影响,Nav1.8被认为是伤害感受的关键参与者。因此,我们发现蛙毒素对该通道具有显著的调节作用:失活过程发生严重改变,激活的电压依赖性向更负的膜电位偏移,导致 Nav1.8 在更负的膜电位下开放,并且离子选择性也发生改变。 蛙毒素 (BTX) 来自南美洲叶蛙属 (Phyllobates) 蛙类,可不可逆地激活电压门控钠通道。先前的研究表明,位于孔道内衬跨膜区段 IVS6 中部附近的苯丙氨酸残基是决定通道对 BTX 敏感性的关键因素。在本研究中,我们在 Na(v)1.3 钠通道的该位点引入了一系列突变,在非洲爪蟾卵母细胞中表达了野生型和突变型通道,并使用电压钳记录技术检测了它们对 BTX 的敏感性。我们发现,丙氨酸或异亮氨酸的取代显著降低了通道对毒素的敏感性,而半胱氨酸、酪氨酸或色氨酸的取代仅轻微降低了毒素的作用。这些数据表明,该位点存在静电配体-受体相互作用,可能涉及BTX上的带电叔胺。随后,我们使用一种对毒素敏感性中等的突变通道(突变体F1710C)来更详细地研究毒素-受体反应的特性。与几乎不可逆地结合BTX的野生型通道不同,毒素从突变通道上的解离速度很快,但仅在通道开放时发生,而非在通道关闭时。这些数据表明,关闭的激活门捕获了结合的毒素。尽管BTX的解离需要通道激活,但矛盾的是,强烈的膜去极化会减缓解离速度,这表明毒素结合反应还受到其他状态依赖性和/或静电因素的影响。我们提出,BTX 通过开放的离子通道孔的胞质端与其受体进行转运,其方式类似于季铵盐类局部麻醉剂(如 QX314)。 我们使用光反应性放射性标记的巴曲毒素衍生物对纯化和重组的大鼠脑钠离子通道进行共价标记,从而定位了巴曲毒素(一种作用于钠离子通道受体 2 位点的脂溶性神经毒素)的结合位点。在短尾拟杆菌(Ptychodiscus brevis)的短尾毒素 1 和拟除虫菊酯类杀虫剂 RU51049(巴曲毒素结合的正向变构增强剂)存在的情况下,一种表观分子量为 240 kDa 的蛋白质(对应于钠离子通道 α 亚基)被特异性共价标记。通过抗体定位蛋白水解片段,鉴定了被光反应性巴曲毒素衍生物特异性光标记的 α 亚基区域。即使经过充分的胰蛋白酶消化,识别钠离子通道跨膜区段IS6邻近氨基酸序列的抗肽抗体仍能免疫沉淀高达70%的标记肽。用胰蛋白酶或V8蛋白酶进行更彻底的消化分析表明,箭毒蛙毒素受体位点部分由结构域I的一部分构成。鉴定出一种特异性免疫沉淀的光标记7.3 kDa肽,该肽包含结构域I的跨膜区段S6,从而将标记位点限定在Asn-388至Glu-429残基(如果V8蛋白酶消化完全)或Leu-380至Glu-429残基(如果消化不完全)。这些结果表明,钠离子通道α亚基I结构域的S6跨膜区是箭毒蛙毒素受体位点的重要组成部分。 甾体神经毒素(-)-箭毒蛙毒素是电压门控钠离子通道(NaV)的强效激动剂。本文报道了箭毒蛙毒素天然(-)和非天然(+)对映体的简洁不对称合成,以及C-20苯甲酸酯修饰衍生物的两种对映异构体的合成。对这些分子针对NaV亚型的电生理特性进行表征,结果表明非天然毒素对映异构体是通道功能的可逆拮抗剂,其活性与(-)-箭毒蛙毒素显著不同。蛋白质诱变实验表明,这些对映异构体在NaV内孔腔中具有共同的结合位点。这些发现激励并推动了后续研究,旨在揭示靶向通道内孔的小分子如何调节钠离子通道(NaV)的动力学。 蛙毒素(BTX):神经毒素,可激活钠离子通道;与洋地黄毒素(地高辛)类似的心脏毒素,可引起心房颤动和心脏骤停。/摘自表格/ |
| 分子式 |
C31H42N2O6
|
|---|---|
| 分子量 |
538.67
|
| 精确质量 |
538.304
|
| CAS号 |
23509-16-2
|
| PubChem CID |
6324647
|
| 外观&性状 |
Noncrystal
|
| 密度 |
1.34g/cm3
|
| 沸点 |
744ºC at 760mmHg
|
| 闪点 |
403.8ºC
|
| 蒸汽压 |
3E-23mmHg at 25°C
|
| 折射率 |
1.642
|
| LogP |
3.49
|
| tPSA |
104.25
|
| 氢键供体(HBD)数目 |
3
|
| 氢键受体(HBA)数目 |
7
|
| 可旋转键数目(RBC) |
4
|
| 重原子数目 |
39
|
| 分子复杂度/Complexity |
1140
|
| 定义原子立体中心数目 |
8
|
| SMILES |
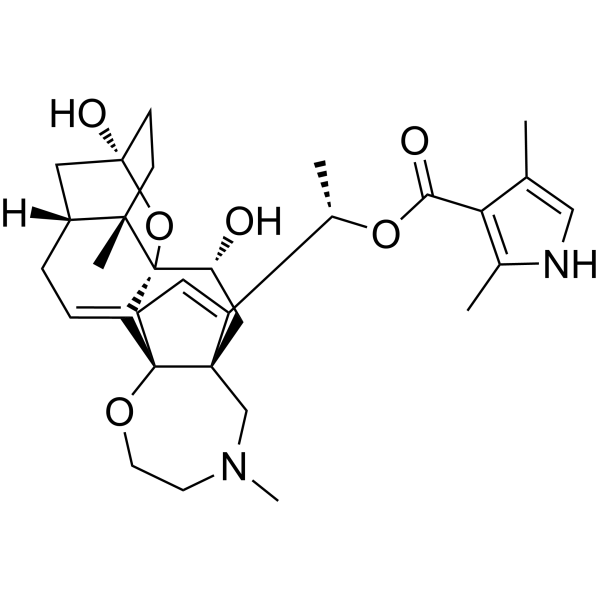
CC1=CNC(=C1C(=O)O[C@@H](C)C2=CC[C@@]34[C@@]2(C[C@H]([C@@]56C3=CC[C@H]7[C@@]5(CC[C@](C7)(O6)O)C)O)CN(CCO4)C)C
|
| InChi Key |
ISNYUQWBWALXEY-OMIQOYQYSA-N
|
| InChi Code |
InChI=1S/C31H42N2O6/c1-18-16-32-19(2)25(18)26(35)38-20(3)22-8-9-30-23-7-6-21-14-29(36)11-10-27(21,4)31(23,39-29)24(34)15-28(22,30)17-33(5)12-13-37-30/h7-8,16,20-21,24,32,34,36H,6,9-15,17H2,1-5H3/t20-,21+,24+,27-,28-,29+,30-,31-/m0/s1
|
| 化学名 |
[(1S)-1-[(1R,5R,6S,9R,11S,12R,14R)-9,12-dihydroxy-6,16-dimethyl-10,19-dioxa-16-azahexacyclo[12.5.3.15,9.01,14.02,11.06,11]tricosa-2,21-dien-22-yl]ethyl] 2,4-dimethyl-1H-pyrrole-3-carboxylate
|
| 别名 |
BTX
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
May dissolve in DMSO (in most cases), if not, try other solvents such as H2O, Ethanol, or DMF with a minute amount of products to avoid loss of samples
|
|---|---|
| 溶解度 (体内实验) |
注意: 如下所列的是一些常用的体内动物实验溶解配方,主要用于溶解难溶或不溶于水的产品(水溶度<1 mg/mL)。 建议您先取少量样品进行尝试,如该配方可行,再根据实验需求增加样品量。
注射用配方
注射用配方1: DMSO : Tween 80: Saline = 10 : 5 : 85 (如: 100 μL DMSO → 50 μL Tween 80 → 850 μL Saline)(IP/IV/IM/SC等) *生理盐水/Saline的制备:将0.9g氯化钠/NaCl溶解在100 mL ddH ₂ O中,得到澄清溶液。 注射用配方 2: DMSO : PEG300 :Tween 80 : Saline = 10 : 40 : 5 : 45 (如: 100 μL DMSO → 400 μL PEG300 → 50 μL Tween 80 → 450 μL Saline) 注射用配方 3: DMSO : Corn oil = 10 : 90 (如: 100 μL DMSO → 900 μL Corn oil) 示例: 以注射用配方 3 (DMSO : Corn oil = 10 : 90) 为例说明, 如果要配制 1 mL 2.5 mg/mL的工作液, 您可以取 100 μL 25 mg/mL 澄清的 DMSO 储备液,加到 900 μL Corn oil/玉米油中, 混合均匀。 View More
注射用配方 4: DMSO : 20% SBE-β-CD in Saline = 10 : 90 [如:100 μL DMSO → 900 μL (20% SBE-β-CD in Saline)] 口服配方
口服配方 1: 悬浮于0.5% CMC Na (羧甲基纤维素钠) 口服配方 2: 悬浮于0.5% Carboxymethyl cellulose (羧甲基纤维素) 示例: 以口服配方 1 (悬浮于 0.5% CMC Na)为例说明, 如果要配制 100 mL 2.5 mg/mL 的工作液, 您可以先取0.5g CMC Na并将其溶解于100mL ddH2O中,得到0.5%CMC-Na澄清溶液;然后将250 mg待测化合物加到100 mL前述 0.5%CMC Na溶液中,得到悬浮液。 View More
口服配方 3: 溶解于 PEG400 (聚乙二醇400) 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 1.8564 mL | 9.2821 mL | 18.5642 mL | |
| 5 mM | 0.3713 mL | 1.8564 mL | 3.7128 mL | |
| 10 mM | 0.1856 mL | 0.9282 mL | 1.8564 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
 Sodium Channel inhibitor 6
Sodium Channel inhibitor 6
 Suzetrigine N-oxide
Suzetrigine N-oxide
 Sodium Channel-IN-8
Sodium Channel-IN-8
 Nav1.2-IN-1
Nav1.2-IN-1
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 463611831
463611831
