| 规格 | 价格 | |
|---|---|---|
| 500mg | ||
| 1g | ||
| Other Sizes |
| 靶点 |
TLR4
|
|---|---|
| 体外研究 (In Vitro) |
微环境启动MSCs导致产生独特的外泌体mRNA含量[4]
当检测未引物MSCs和CRX-527引物MSCs之间的外泌体mrna类型时,1915个蛋白质编码基因在CRX-527处理后表现出差异表达(FDR < 5%, >是标准化读取计数的2倍变化)。在CRX-527诱导的上调外泌体mrna中,“炎症反应”基因本体(GO)项(GO:0002544)显著富集(图2;调整p值= 3.8E−4)。除了炎症反应GO项外,外泌体mrna的上调还增强了多种生物学功能,如成纤维细胞迁移的正调节(GO:0010763;调整p值= 6.2 2e−12),内在凋亡信号通路的调节(GO:2001242;调整后的p值= 1.4E−7),纤溶酶原活化(GO:0031639;调整p值= 8.9E−7)。这些结果表明,CRX-527的启动改变了MSCs外泌体mrna的含量,表明体外微环境刺激在调节炎症和伤口愈合中的作用[4]。 为了确定外泌体mRNA含量的变化是对特定微环境刺激的特异性还是对MSC启动的一般反应,我们比较了两种不同的MSCs启动方法:TNF-α和CRX-527对外泌体mRNA含量变化重叠程度的影响。我们观察到两种MSC启动方法之间差异表达的外泌体mrna存在适度重叠(图3)。例如,在响应CRX-527启动的180个外泌体mrna中,只有11个(6.1%)响应TNF-α启动而上调。至于下调的基因,受CRX-527启动的MSCs影响的1735个外泌体mrna中有857个(49.4%)与受TNF-α启动的MSCs影响的外泌体重叠。此外,与每种MSC启动方法相关的mRNA含量变化被发现富集在不同的生物学功能中(图3)。例如,血管内皮细胞迁移和成纤维细胞迁移项的正调节特异性富集在CRX-527启动的MSC-exosomes上调mRNA中,而不富集在TNF-α启动的MSC-exosomes上调mRNA中。这一观察结果表明,外泌体mRNA含量受MSCs特定微环境的控制,在TNF-α和TLR4启动后存在差异。 MSCs的微环境启动导致外泌体microRNA含量的调节[4] 最近的证据表明,外泌体的microRNA装载是一个选择性和受调节的过程,而不是随机发生的。我们试图确定MSC微环境启动的类型是否影响外泌体的microRNA含量。为了实现这一目标,我们进行了小RNA测序,以分析来自天然MSCs、CRX-527引物MSCs和TNF-α引物MSCs的外泌体中的短RNA片段。我们分别鉴定了31个响应CRX-527治疗的差异表达外泌体microrna(图4A)和23个响应TNF-α治疗的差异表达外泌体microrna(图4B)。值得注意的是,在CRX-527和TNF-α处理之间,没有差异表达的外泌体microRNA重叠,这表明外泌体microRNA含量受到用于启动MSCs的特定微环境刺激的影响。根据TNF-α引物的msc -外泌体microRNA靶点的富集p值,“巨噬细胞分化”是富集程度最高的基因本体(GO)术语(图4D)。TNF-α是一种促炎细胞因子,已知可诱导巨噬细胞的m1样极化。丰富“巨噬细胞分化”GO术语表明,作为分子货物,外泌体可能在促炎刺激(如TNF-α扰动)下运输特定的microrna,启动单核细胞分化为巨噬细胞。图4C和D分别显示了CRX-527和TNF-α启动的最显著p值,显示了前3个富集的氧化石墨烯项。 亲本细胞与外泌体之间RNA丰度的相关性[4] 我们进一步对MSCs进行了mRNA和小RNA测序,以研究亲代MSCs与其外泌体之间的RNA丰度是否相关。我们观察到外泌体与其亲本细胞在天然条件下的适度相关性,microrna和mrna的Spearman 's Rank相关系数分别为0.46和0.57(图5)。这表明,一部分外泌体RNA(但不是全部)确实再现了它们亲本间质干细胞的RNA丰度。当MSCs暴露于微环境刺激(如CRX-527和TNF-α启动)时,这些相关性增加,尤其是microRNA(图5)。外泌体和亲本MSC microRNA丰度之间的Spearman等级相关系数(Rho)分别从0.46(在天然条件下)增加到0.78和0.76,CRX-527和TNF-α启动的MSCs。这些数据表明,MSCs的微环境刺激使MSCs的microRNA谱与其分泌的外泌体同步。mrna也出现了类似的趋势,尽管不那么明显(图5)。因此,msc -外泌体rna不仅仅是来自亲本细胞的随机rna子集,而是受驱动外泌体生成的微环境刺激的调控和依赖。 |
| 体内研究 (In Vivo) |
Morroniside(Mor)是山茱萸中的一种生物活性化合物,具有抗炎、神经保护和抗氧化特性。长期使用麻醉剂七氟烷(Sev)与术后认知功能障碍(POCD)的发展有关。本研究旨在阐明Mor改善认知障碍的作用机制。在老年小鼠中建立了Sev诱导的认知功能障碍模型,并使用水迷宫实验进行了行为分析。通过苏木精和伊红(HE)染色、尼氏染色和TUNEL染色观察小鼠海马的组织病理学变化和神经元凋亡。ELISA和qRT-PCR检测炎症因子水平。通过免疫荧光、流式细胞术和qRT-PCR评估海马组织中小胶质细胞的表型转化。使用分子对接分析Mor和TLR4之间的相互作用。Western blot鉴定了凋亡相关蛋白、突触相关蛋白和TLR4/NF-κB通路蛋白的水平。吸入Sev可显著降低老年小鼠的学习和空间记忆能力,Mor可剂量依赖性地改善这一现象。Mor抑制神经炎症,调节海马小胶质细胞的极化状态,促进其极化为M2型,减轻Sev诱导的海马组织损伤和神经元凋亡。值得注意的是,Mor可以很好地与TLR4结合,并减少TLR4的阳性表达。Mor阻断了Sev诱导的海马组织TLR4/NF-κB通路激活,TLR4激动剂CRX-527减弱了Mor的作用。总之,Mor阻断了TLR4/NF-κB通路,减少了Sev引起的海马组织损伤和神经炎症,从而改善了老年小鼠的认知障碍[1]。
近年来,toll样受体(Toll-like receptor, TLRs)在辐射损伤中的研究得到了广泛的研究,但大多数TLR配体的高毒性和低疗效的固有缺陷限制了其进一步的临床转化。CRX-527作为TLR4的一种配体,对辐射的保护作用很少有报道。实验证明,在相同剂量下,CRX-527在体内比LPS更安全,在体外几乎没有毒性作用。给药CRX-527可使野生型小鼠全身照射(TBI)的存活率提高到100%,而TLR4-/-小鼠则无此作用。TBI后,crx -527治疗小鼠造血系统损伤明显减轻,恢复期加快。此外,CRX-527诱导hsc分化,刺激CRX-527可显著增加LSK细胞的比例和数量,促进其向巨噬细胞分化,激活免疫防御。此外,我们提出造血分化在保护肠道免受辐射损伤中的免疫防御作用,并证实巨噬细胞通过外周血侵入肠道,保护肠道免受辐射损伤。同时,CRX-527维持肠道功能和稳态,促进肠道干细胞再生,保护肠道免受致死剂量辐照损伤。此外,在小鼠使用后,我们发现CRX-527对辐照后的TLR4-/-小鼠的造血系统和肠道系统没有明显的保护作用。综上所述,CRX-527诱导造血干细胞分化,保护肠上皮免受辐射损伤 TLR4在VD小鼠中介导NTP抗炎作用[3] 鉴于NTP的神经保护作用可能至少部分由TLR4介导,我们假设TLR4拮抗剂在抑制炎症和改善记忆方面可能具有与NTP相似的作用,而TLR4激动剂可能逆转NTP的保护作用。为了进一步阐明NTP的抗炎作用是否至少部分由TLR4介导,我们使用了TLR4特异性拮抗剂TAK242和特异性激动剂CRX-527。将小鼠随机分为假手术组、BCAS/生理盐水(NS)组、BCAS/NTP组、BCAS/TAK242组和BCAS/NTP/CRX-527组,每组12只。[3] 我们发现,NTP或TAK-242治疗后,IL-1β、IL -6和TNFα mRNA水平降低。然而,NTP与CRX-527联合使用导致这些mRNA水平升高(IL-1β: P<0.001;il - 6: P < 0.001;TNFα: P<0.001,与BCAS/NTP组相比)(图7A,B和7C)。同样,给药后MyD88和pP65蛋白水平显著升高(MyD88: P<0.001;pP65: P<0.001,与BCAS/NTP组相比)(图7E和F),提示NTP在VD小鼠中至少部分通过TLR4/MyD88/NF-κB途径发挥其抗炎作用。[3] 接下来,我们观察了这两种抑制剂对神经胶质细胞的影响。BCAS/NTP/CRX-527组海马CA1区和CA3区IBA-1阳性细胞多于BCAS/NTP组(CA1: P<0.001;CA3: P = 0.004)(图6A和B)。此外,GFAP荧光染色显示,与BCAS/NTP/CRX-527组相比,BCAS/NTP组CA1和CA3区阳性细胞数量显著增加(CA1: P = 0.0037;CA3: P<0.001)(图6C和D)。 为了评估BCAS/NTP组小鼠记忆功能的改善是否与TLR4相关,小鼠在治疗后进行y迷宫测试。与假手术组相比,BCAS/NS组Y-maze自发交替率显著降低(F(4,47) = 27.15,P = 0.000)。与BCAS/NS组相比,NTP和TAK242治疗的正确率显著增加。然而,与BCAS/NTP组相比,CRX-527明显消除了NTP对BCAS小鼠%正确交替的治疗作用(P<0.001)。此外,BCAS/NTP和BCAS/TAK242组之间没有差异(P>0.5)(图8),这表明TLR4的过度激活逆转了NTP对记忆损伤的影响,并且TLR4进一步介导了NTP对VD小鼠记忆的改善。 |
| 细胞实验 |
共培养试验[2]
简单地说,1×105 RAW264.7或THP-1细胞在Transwell室(BIOFIL, 0.4µm, 6.5 mm直径)中接种,MODE-K或hiec细胞在12孔板底部接种,然后根据制造商的说明进行培养。RAW264.7细胞或Transwell室THP-1细胞在辐照前12小时用CRX-527处理。照射后,使用12孔板上的MODE-K细胞或hiec进行菌落形成、ROS和Western blot分析。 损伤韧带的处理包括:(1)PBS(作为损伤对照),(2)5 × 106外泌体(exosome), (3) TNF-α-引物MSCs (TNF)的5 × 106外泌体,或(4)CRX-527-引物MSCs (CRX)的5 × 106外泌体。外泌体剂量的选择基于先前的结果,结果显示1 × 106外泌体的递送降低了M1/M2巨噬细胞比例,但没有改善肌腱功能。5 × 106外泌体的剂量是撕裂MCL中可以添加的最大剂量和体积,而不会出现过多的渗漏。给药后缝合皮肤。在损伤后3天(当巨噬细胞的存在升高时),通过针和注射器将第二剂量的5 × 106外泌体施用于对侧MCL,以检查额外剂量的外泌体是否可以提供对愈合的附加效应。伤后14天,收集mcl进行力学测试。由于没有观察到力学结果中的依赖效应,因此将两方合并进行所有数据分析。[4] |
| 动物实验 |
雄性SPF级C57BL/6J小鼠(20月龄)饲养于洁净级动物房内。室温23℃~28℃,湿度45%~55%,12小时光照/12小时黑暗循环,小鼠自主进食饮水。动物实验遵循3R原则,每日更换垫料,并定期消毒饲料容器、笼子、水瓶、饮水管等设施。适应性喂养一周后,将小鼠随机分为对照组(n=8)、Sev组(n=8)、Sev-Mor组(n=16)和Sev-Mor-TLR4激动剂组(n=4)。使用麻醉气体监测仪监测Sev、二氧化碳和氧气浓度。在诱导箱底部铺撒少量碱石灰以防止二氧化碳积聚。为防止小鼠吸入钠石灰粉尘导致灼伤,将透气隔离垫平铺在钠石灰上。根据以下方法建立了七氟醚麻醉下认知功能障碍的小鼠模型。
小鼠持续吸入2%七氟醚5小时,总气流为1.5L/min,载气为70%氧气。七氟醚麻醉结束后,将小鼠唤醒并放回干燥温暖的笼子中。对照组:正常吸入室内空气。对照组小鼠腹腔注射生理盐水(0.1 mL/100 g)。七氟醚组小鼠麻醉后腹腔注射生理盐水。 S-Mor 组小鼠在接受 Mor(30、60 和 100 mg/kg 体重)治疗前进行麻醉,每三天一次,持续四周。S-Mor-CRX-527 组小鼠在麻醉后,每三天腹腔注射 Mor(100 mg/kg 体重),连续四周,然后在 Mor 治疗的最后三天,每天腹腔注射一次 CRX-527(0.25 mg/kg 体重,V42156,InvivoChem LLC)。 [1] 照射和治疗[2] 使用照射器(KUBTEC XCELL 225,225 kV,13.2 mA,1 Gy/min)对HIECs和MODE-K细胞进行单次16 Gy X射线照射,同时在相同条件下平行研究未照射的对照细胞。对于小鼠,采用5 Gy全身剂量观察造血系统的变化,采用7.5 Gy全身剂量观察肠道系统的变化,并采用9 Gy局部腹部照射观察肠道系统的变化。小鼠在照射前24小时和2小时腹腔注射0.5 mg/kg的CRX-527。全身照射的小鼠用固定架固定后,放入照射器中进行照射。照射后,收集相应的组织进行检测。 VD小鼠模型的建立及药物治疗[3] 我们采用双侧颈总动脉模型(BCAS)建立了VD小鼠模型(Shibata等,2004;Shibata等,2007;Ihara等,2014)。适应两周后,所有小鼠随机分为五组:假手术组、BCAS/生理盐水(NS)组、BCAS/NTP组、BCAS/TAK242组和BCAS/NTP/CRX-527组(每组n=12)。具体而言,小鼠用3.0%异氟烷麻醉,并仰卧放置。为了暴露双侧颈总动脉,在颈部正中线做一个小切口。将直径 0.18 mm、长度 2.5 mm 的微线圈分别置入双侧颈总动脉。先在一侧放置线圈,约一小时后放置另一侧。术中及术后体温均维持在 36.5 ± 0.5 °C,使用加热垫进行加热。小鼠麻醉苏醒后,即可在笼内自由活动,并可自由获取水和食物。术后三天,BCAS/NS组和BCAS/NTP组分别给予生理盐水和NTP(50NU/Kg,每日一次),持续28天(Fang等,2019)。为了观察NTP对TLR4的影响,根据之前的研究和制造商的说明,腹腔注射3 mg/kg TAK-242或0.25 mg/kg CRX-527(tlrl-crx527)(Hua等,2015;Yang等,2020;Zhang等,2022)。 |
| 参考文献 |
[1]. Morroniside ameliorates sevoflurane anesthesia-induced cognitive dysfunction in aged mice through modulating the TLR4/NF-κB pathway. Biomol Biomed. 2024 Dec 6. doi: 10.17305/bb.2024.11433.
[2]. CRX-527 induced differentiation of HSCs protecting the intestinal epithelium from radiation damage. Front Immunol. 2022 Aug 30:13:927213. [3]. Neurotropin alleviates cognitive impairment by inhibiting TLR4/MyD88/NF-κB inflammation signaling pathway in mice with vascular dementia. Neurochem Int. 2023 Dec:171:105625. [4]. Modulating the mesenchymal stromal cell microenvironment alters exosome RNA content and ligament healing capacity. Stem Cells. 2024 Jul 8;42(7):636-649. |
| 其他信息 |
总之,我们的研究首次证实,Mor能够减轻老年小鼠海马中Sev诱导的组织病理损伤,促进小胶质细胞向M2型转化,并抑制神经炎症。重要的是,Mor能够阻断TLR4/NF-κB通路,提示其可能通过调节该通路来缓解老年小鼠的认知功能障碍。本研究阐明了Mor缓解老年小鼠认知功能障碍的潜在作用机制,为术后认知功能障碍(POCD)的临床治疗提供了新的参考。然而,本研究仍存在一些不足。由于时间和条件的限制,本研究的样本量较小。未来需要进一步探索Mor对幼鼠认知功能障碍的影响及其作用机制。 [1] 本研究表明,CRX-527 可保护机体免受辐射引起的造血和肠道损伤。与 LPS 相比,CRX-527 在体内和体外的毒性均较低。机制上,CRX-527 的刺激显著增加了造血干细胞 (HSC) 的比例和数量,并促进其分化为巨噬细胞,从而激活免疫防御。在此基础上,我们观察到电离辐射后小鼠肠道结构和功能的积极变化。TLR4 相关通路的激活介导了 CRX-527 对造血和肠道辐射损伤的保护作用。[2] 在本研究中,TAK242 抑制了 BCAS 诱导的神经炎症,其作用与 NTP 类似。最后,我们给予 TLR4 特异性激动剂 CRX-527 以阐明 NTP 作用的潜在机制。 TLR4特异性激动剂CRX-527在刺激后24小时显著激活了NF-κB启动子(Bowen等,2012)。CRX-527诱导下游MyD88信号级联的激活,导致NF-κB的早期激活和核转位,从而促进炎症(Zhang等,2022)。我们发现CRX-527消除了NTP的作用,加重了神经炎症,并增加了记忆障碍,表明TLR4在VD炎症中起着重要作用。NTP在VD小鼠中的抗炎作用是通过调节TLR4实现的。这提示NTP可能阻断了神经通路中传感器分子的激活,从而为神经炎症性疾病的治疗提供新的思路。综上所述,我们发现NTP通过TLR4/MyD88/NF-κB通路抑制炎症。 [3]
尽管间充质干细胞(MSC)疗法在再生医学领域展现出巨大潜力,但由于免疫相容性等问题,其临床应用仍面临挑战。MSC来源的外泌体是一种很有前景的即用型疗法,能够以无细胞的方式促进伤口愈合。然而,MSC来源外泌体的内容物定制化及其对组织再生作用的影响机制仍有待深入研究。本研究采用体外系统,比较了两种炎症诱导剂TNF-α和CRX-527(一种高效的合成TLR4激动剂,可用作疫苗佐剂或诱导抗肿瘤免疫)对人MSC启动作用及其对外泌体分子载荷的影响,并利用体内大鼠韧带损伤模型验证了外泌体的效力。体外培养间充质干细胞(MSCs)时,不同的微环境刺激会影响其外泌体中的microRNA和mRNA,进而影响韧带愈合。与炎症诱导剂处理的MSCs来源的外泌体相比,未经处理的MSCs来源的外泌体能显著增强愈合韧带的力学性能;而炎症诱导剂处理的MSCs来源的外泌体不仅未能改善韧带的力学性能,反而可能使其力学性能恶化。此外,研究还发现外泌体microRNA水平的改变与韧带中microRNA靶基因的表达变化之间存在关联。这些发现阐明了MSCs、其外泌体和组织再生之间错综复杂的相互作用。[4] |
| 分子式 |
C81N2O19PH151
|
|---|---|
| 分子量 |
1488.041
|
| 精确质量 |
1,487.06
|
| 元素分析 |
C, 65.38; H, 10.23; N, 1.88; O, 20.43; P, 2.08
|
| CAS号 |
216014-14-1
|
| 相关CAS号 |
216014-14-1 (CRX-527);216014-05-0 (CRX-547);
|
| 外观&性状 |
Typically exists as solid at room temperature
|
| SMILES |
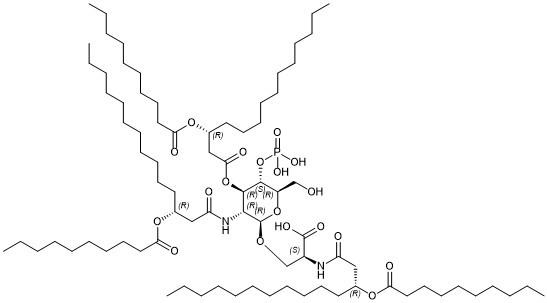
O=C([C@@H](NC(C[C@@H](CCCCCCCCCCC)OC(CCCCCCCCC)=O)=O)CO[C@H]1[C@@H]([C@H]([C@@H]([C@H](O1)CO)OP(O)(O)=O)OC(C[C@@H](CCCCCCCCCCC)OC(CCCCCCCCC)=O)=O)NC(C[C@@H](CCCCCCCCCCC)OC(CCCCCCCCC)=O)=O)O
|
| InChi Key |
REEGNIYAMZUTIO-MGSMBCBTSA-N
|
| InChi Code |
InChI=1S/C81H151N2O19P/c1-7-13-19-25-31-34-40-43-49-55-66(97-73(87)58-52-46-37-28-22-16-10-4)61-71(85)82-69(80(91)92)65-96-81-77(83-72(86)62-67(56-50-44-41-35-32-26-20-14-8-2)98-74(88)59-53-47-38-29-23-17-11-5)79(78(70(64-84)100-81)102-103(93,94)95)101-76(90)63-68(57-51-45-42-36-33-27-21-15-9-3)99-75(89)60-54-48-39-30-24-18-12-6/h66-70,77-79,81,84H,7-65H2,1-6H3,(H,82,85)(H,83,86)(H,91,92)(H2,93,94,95)/t66-,67-,68-,69+,70-,77-,78-,79-,81-/m1/s1
|
| 化学名 |
O-((2R,3R,4R,5S,6R)-3-((R)-3-(decanoyloxy)tetradecanamido)-4-(((R)-3-(decanoyloxy)tetradecanoyl)oxy)-6-(hydroxymethyl)-5-(phosphonooxy)tetrahydro-2H-pyran-2-yl)-N-((R)-3-(decanoyloxy)tetradecanoyl)-L-serinate
|
| 别名 |
CRX-527; 216014-14-1; Decanoic acid, (1R)-1-[2-[[(1S)-1-carboxy-2-[[2-deoxy-3-O-[(3R)-1-oxo-3-[(1-oxodecyl)oxy]tetradecyl]-2-[[(3R)-1-oxo-3-[(1-oxodecyl)oxy]tetradecyl]amino]-4-O-phosphono-beta-D-glucopyranosyl]oxy]ethyl]amino]-2-oxoethyl]dodecyl ester
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
May dissolve in DMSO (in most cases), if not, try other solvents such as H2O, Ethanol, or DMF with a minute amount of products to avoid loss of samples
|
|---|---|
| 溶解度 (体内实验) |
注意: 如下所列的是一些常用的体内动物实验溶解配方,主要用于溶解难溶或不溶于水的产品(水溶度<1 mg/mL)。 建议您先取少量样品进行尝试,如该配方可行,再根据实验需求增加样品量。
注射用配方
注射用配方1: DMSO : Tween 80: Saline = 10 : 5 : 85 (如: 100 μL DMSO → 50 μL Tween 80 → 850 μL Saline)(IP/IV/IM/SC等) *生理盐水/Saline的制备:将0.9g氯化钠/NaCl溶解在100 mL ddH ₂ O中,得到澄清溶液。 注射用配方 2: DMSO : PEG300 :Tween 80 : Saline = 10 : 40 : 5 : 45 (如: 100 μL DMSO → 400 μL PEG300 → 50 μL Tween 80 → 450 μL Saline) 注射用配方 3: DMSO : Corn oil = 10 : 90 (如: 100 μL DMSO → 900 μL Corn oil) 示例: 以注射用配方 3 (DMSO : Corn oil = 10 : 90) 为例说明, 如果要配制 1 mL 2.5 mg/mL的工作液, 您可以取 100 μL 25 mg/mL 澄清的 DMSO 储备液,加到 900 μL Corn oil/玉米油中, 混合均匀。 View More
注射用配方 4: DMSO : 20% SBE-β-CD in Saline = 10 : 90 [如:100 μL DMSO → 900 μL (20% SBE-β-CD in Saline)] 口服配方
口服配方 1: 悬浮于0.5% CMC Na (羧甲基纤维素钠) 口服配方 2: 悬浮于0.5% Carboxymethyl cellulose (羧甲基纤维素) 示例: 以口服配方 1 (悬浮于 0.5% CMC Na)为例说明, 如果要配制 100 mL 2.5 mg/mL 的工作液, 您可以先取0.5g CMC Na并将其溶解于100mL ddH2O中,得到0.5%CMC-Na澄清溶液;然后将250 mg待测化合物加到100 mL前述 0.5%CMC Na溶液中,得到悬浮液。 View More
口服配方 3: 溶解于 PEG400 (聚乙二醇400) 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 0.6720 mL | 3.3601 mL | 6.7202 mL | |
| 5 mM | 0.1344 mL | 0.6720 mL | 1.3440 mL | |
| 10 mM | 0.0672 mL | 0.3360 mL | 0.6720 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
 匹伐加宾
匹伐加宾
 磷酸氢化可的松
磷酸氢化可的松
 BAY-784
BAY-784
 Gly-PEG3-endo-BCN
Gly-PEG3-endo-BCN
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1
































 463611831
463611831
