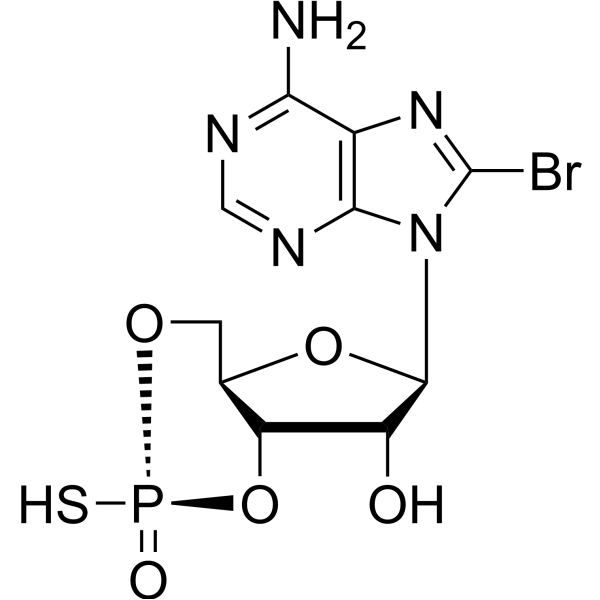
| 规格 | 价格 | |
|---|---|---|
| 500mg | ||
| 1g | ||
| Other Sizes |

| 靶点 |
PKA (EC50 = 360 nM); cAMP analog
|
|---|---|
| 体外研究 (In Vitro) |
Sp-8-Br-cAMPS (0-100 nM) 可抑制嗜线虫病菌 (X. nematophila) 和枯草芽孢杆菌 (B. subtilis) 的吞噬作用,但不影响每种血细胞类型的细菌数量[1]。Sp-8-Br-cAMPS (0-1000 μM) 剂量依赖性地通过激活 PKA 抑制葡萄球菌肠毒素 B (SEB) 诱导的 T 细胞活化和细胞因子 IFN-γ、TNF-α、IL-2 和 IL-4 的表达[2]。
PKA激活剂以浓度依赖的方式严重损害了嗜线虫X.的吞噬作用(图3a),而枯草芽孢杆菌的抑制作用仅在高浓度下发生(图3b)。附着任何一种细菌的颗粒细胞水平也降低了。尽管嗜线虫X.的浆细胞水平下降,但枯草芽孢杆菌的水平没有变化。酶激活剂不影响每种血细胞类型的细菌数量(X.嗜线虫:对照,1.7±0.3个细菌/浆细胞,2.1±0.3个菌/颗粒细胞;100 nmol/LSp-8-Br-cAMPS,1.3±0.2个细菌/血浆细胞,1.9±0.3个杆菌/颗粒细胞[P>0.05];枯草芽孢杆菌:对照,2.0±0.4个细菌/血细胞,1.7±0.3/颗粒细胞,100 nmol/L Sp-8-Br cAMPS,1.4±0.3个菌株/浆细胞、1.2±0.2个菌/粒细胞[P>0.05])。然而,检测到每种血细胞类型的细菌数量持续下降。将血细胞与50 nmol/L PKA激活剂和越来越多的PKA抑制剂共同孵育,可以提高两种细菌的颗粒细胞和浆细胞水平(图4)。[1] PKA调节剂本身会影响血细胞蛋白的释放。酶抑制剂浓度的增加会增加蛋白质的释放,而激活剂浓度的增加则会损害蛋白质的释放(图5b)。Rp-8-Br-cAMPS抵消了PKA激活剂Sp-8-Br-McAMPS对蛋白质释放的抑制。[1] cAMP和PKA I型抑制SEB诱导的T细胞活化和IFN-γ的表达[2] 为了探索cAMP和PKA I型对免疫激活早期功能事件的可能影响,我们在用超抗原SEB激活PBMC 6小时后测量了激活标志物CD69的表达和IFN-γ的产生。在PKA激动剂单独存在或不存在的情况下,通过流式细胞术测量CD69和IFN-α的表达。将细胞与cAMP类似物预孵育1小时,以使化合物扩散,然后用SEB激活。然后在用SEB激活后1小时加入布雷菲尔丁A,并将培养物孵育5小时以积累细胞因子。 与对照组相比,CD3+T细胞中CD69和IFN-γ的表达在与250μMSp-8-Br-cAMPS(激动剂)预孵育后显著降低(图1,a和b)。为了阐明这种抑制的细胞因子产生是通过与TCR/CD3和脂筏相关的PKA I型还是PKA II型介导的,PKA II的定位主要局限于淋巴细胞的核周和高尔基中心体区域(17),我们用250μM Sp-8-Br-cAMPS与1000μM PKA I类选择性拮抗剂Rp-8-Br-cAMPS联合预孵育细胞(图1,c)。拮抗剂几乎完全逆转了激动剂的抑制作用,表明cAMP在早期T细胞活化中的抑制作用主要是由于PKA I型的活化。拮抗剂单独对CD69和IFN-γ的表达没有影响(图1d)。 cAMP和PKA I型的抑制作用不是由于细胞凋亡[2] 为了研究cAMP和PKA I型的抑制作用是否是由于诱导凋亡引起的,我们通过流式细胞术在正向和侧向散射上评估了未刺激和SEB刺激的PBMC在与1000μM Sp-8-Br-cAMPS预孵育和未与布雷菲丁A预孵育的最后5小时的形态。将培养物孵育6至48小时的不同时间段,在cAMP类似物存在的情况下,没有表现出凋亡的形态学变化特征,前向散射减少,侧向散射增加(数据未显示)。此外,在相同的实验条件下,还通过Annexin VFITC标记评估了细胞凋亡(图2显示了培养24小时)。cAMP激动剂在静息(图2,a和b)或活化培养物(图2、c和d)中没有诱导膜联蛋白V结合作为凋亡细胞标志的任何显著增加。相比之下,用作对照的茴香霉素(图2,e和f)在未刺激和活化的培养物中诱导了细胞凋亡并增加了膜联蛋白V的结合。通过碘化丙啶排除法获得了类似的数据(数据未显示)。 cAMP和PKA I型抑制SEB诱导的IFN-γ、TNF-α、IL-2和IL-4[2] 为了检查cAMP和PKA I型对IFN-γ表达的影响是否是特异性的,我们测量了在SEB激活PBMC后,随着Sp-8-Br-cAMPS浓度的增加,各种细胞因子的表达。我们发现,cAMP类似物以浓度依赖的方式降低了所有检测到的细胞因子(IFN-γ、TNF-α、IL-2和IL-4)的表达(图3a),尽管cAMP类似物对抑制的敏感性因细胞因子而异。在所有情况下,cAMP介导的抑制作用可被PKA I型选择性拮抗剂Rp-8-Br-cAMPS逆转。接下来,我们测量了细胞因子积累期较长时细胞因子表达的抑制水平,以观察这种作用是否是短暂的。进行了5、15和25小时的累积期,在所有这些时间点,抑制水平都是持续的,并且具有浓度依赖性(数据未显示)。 cAMP和PKA I型独立于APC抑制CD4和CD8 T细胞活化[2] 为了进一步解决cAMP在抑制免疫激活中影响哪个T细胞室的问题,我们研究了cAMP和PKA I型对CD4和CD8 T细胞的影响。如图5、a和b所示,IFN-γ和TNF-α的表达均以浓度依赖的方式受到抑制。然而,该实验是在含有APC的PBMC的SEB刺激下进行的。为了消除观察到的效应可能是由于cAMP对APC功能的抑制作用造成的可能性,我们通过粘附分离APC,用SEB装载细胞,然后用多聚甲醛固定。然后,我们以1:10的比例建立了与固定的载银APC和APC耗竭的PBMC群体的自体共培养。在加入布雷菲尔丁A之前,将共培养物孵育6小时。允许细胞因子积累14小时。通过细胞内流式细胞术评估CD4和CD8 T细胞中IFN-γ的表达,观察到细胞因子表达的浓度依赖性抑制,随着Sp-8-Br-cAMPS浓度的增加,这表明抑制作用与APC无关(图5c)。 细胞因子表达和增殖的联合抑制[2] 为了进一步确定cAMP介导的免疫功能抑制的程度,我们通过CFSE稀释试验研究了对CD3、CD4和CD8 T细胞增殖的影响。如图6a所示,在所有三个细胞群中,随着Sp-8-Br-cAMPS浓度的增加,SEB诱导的增殖呈浓度依赖性下降。PKA I型选择性拮抗剂Rp-8-Br-cAMPS的抑制是可逆的(数据未显示)。 通过将CFSE稀释试验与细胞内流式细胞术相结合,我们同时测量了cAMP对两种免疫功能的影响。如图6b所示,每个细胞分裂的CFSE荧光强度依次降低,表达IFN-γ的T细胞比例最高的是后代细胞。随着Sp-8-Br-cAMPS浓度的增加,IFN-γ表达显著降低,增殖受到明显抑制,表现为1000μM Sp-8-Br cAMPS的最后两次细胞分裂消失。 cAMP和PKA I型抑制Ag特异性免疫反应[2] 接下来,我们研究了cAMP和PKA I型对Ag特异性免疫反应的影响(图7a)。如SEB诱导的免疫反应所示,我们观察到,通过增加CD4和CD8 T细胞群中Sp-8-Br-cAMPS的浓度,CMV诱导的IFN-γ表达受到浓度依赖性抑制。为了诱导最佳刺激,CD4 T细胞群被CMV感染的细胞提取物刺激,而CD8 T细胞群则被CMVpp65刺激。两种细胞群的抑制范围均为60%至80%。 我们还通过结合CFSE稀释试验和细胞内流式细胞术,研究了PKA I型激活对结核菌素PPD诱导的T细胞增殖和IFN-γ产生的影响(图7b)。正如SEB和CMV诱导的免疫反应所观察到的那样,结核菌素PPD诱导的T细胞增殖和IFN-γ表达以浓度依赖的方式下降,Sp-8-Br-cAMPS。此外,cAMP拮抗剂Rp-8-Br-cAMPS可完全逆转T细胞增殖和IFN-γ表达(数据未显示)。 cAMP激活Csk[2] 为了评估本研究中使用的cAMP激动剂对先前绘制的cAMP-PKA抑制T细胞功能的机制的影响(11),我们检查了Sp-8-Br-cAMPS是否会导致Csk活性的增加。在用cAMP类似物处理浓度增加后,体外评估了Csk磷酸转移酶活性,导致Csk活性呈浓度依赖性增加,使用最高浓度时活性增加了2倍以上(图8)。 |
| 体内研究 (In Vivo) |
Sp-8-Br-cAMPS (50 nM) 可抑制血细胞释放蛋白质,抑制细菌从血淋巴中清除,但不影响 G. mellonella 幼虫的活力 [1]。
单独注射Sp-8-Br-cAMPS后30分钟内,与PBS对照幼虫相比,总血细胞计数增加,而PKA抑制剂降低了血细胞水平(图7)。在含有抑制剂的昆虫中观察到结节(直径1.2±0.1mm)和广泛的血细胞聚集(2×105/mL),但在含有激活剂的组中没有。增加抑制剂的量会降低含有固定量PKA激活剂的幼虫的血细胞计数。 |
| 酶活实验 |
PKA生化测定[1]
为了确定PKA调节剂影响PKA活性以及PKA活性在血细胞粘附过程中发生变化,使用了PKA生化测定。将冷冻幼虫放血,并将180µL血淋巴加入200µL抗凝剂中23。将200微升血细胞悬浮液离心(200 g,2分钟,4°C),通过离心将血细胞沉淀在100µL PBS中洗涤两次。将第一组颗粒重新悬浮在20µL PBS中,PBS中含有选定量的Rp-8-Br-cAMPS和/或Sp-8-Br-cAMPS。孵育30分钟后,在1 mL PBS中离心三次洗涤细胞,通过剧烈移液溶解,并根据制造商的说明测定PKA活性。将第二组血细胞放置在含有玻璃盖玻片(直径5mm)的96孔微量滴定板中。为了防止血细胞粘附引起PKA活性的可能变化,立即裂解血细胞作为非附着血细胞对照组。在室温下孵育30分钟后,其他孔中主要代表粘附血细胞的细胞被裂解。测定两组PKA活性。四个重复中的每一个至少使用两个样品。 Csk活性评估[2] 纯化的外周T细胞(RPMI 1640中为40×106个细胞/ml)在37°C下孵育30分钟,加入或不加入指定浓度的cAMP类似物Sp-8-Br-cAMPS。此后,将细胞在裂解缓冲液(50 mM HEPES(pH 7.4)、100 mM NaCl、5 mM EDTA、1%Triton X-100、50 mM n-β-辛基葡萄糖苷、10 mM NaPPi、1 mM Na3VO4、50 mM NaF和1 mM PMSF)中破碎,并用抗Csk Ab进行免疫沉淀。在4°C下孵育过夜后,加入蛋白A-Sepharose并继续孵育1小时。免疫复合物在裂解缓冲区中洗涤三次,在Csk激酶测定缓冲液(50mM HEPES和5mM MgCl2,pH 7.4)中洗涤三倍,然后进行Csk激酶分析和Western blot分析。人Csk的酪氨酸激酶活性是通过将[32P]磷酸盐掺入合成的聚氨基酸聚(Glu,Tyr)4:1中来测量的。遵循标准方案,反应体积为50μl,含有HEPES缓冲液(pH 7.4)、5 mM MgCl2、200μM[γ-32P]ATP(0.15 Ci/mmol)、200μg/ml聚(Glu,Tyr)和免疫沉淀的Csk。孵育温度为30°C,孵育时间为12分钟。 |
| 细胞实验 |
细菌吞噬作用和血细胞与相关细菌[1]
通过将从总共六只在冰上冷却(15分钟)的幼虫中收集的60µL昆虫血淋巴加入1 mL冰冷的PBS(1.6×106个血细胞/mL)中制备血细胞单层。除非另有说明,否则血细胞不会被洗掉血浆,这样它们更接近体内情况。将15微升悬浮液置于无内毒素载玻片上,在相对湿度>95%的条件下孵育30分钟,使血细胞粘附。通过用2mL PBS冲洗载玻片三次来去除未附着的血细胞。将含有和不含选定浓度PKA调节剂(PKA激活剂Sp-8-Br-cAMPS;PKA I型抑制剂RP-8-Br-cAMPS27)的PBS中的荧光细菌添加到单层中,使细菌与血细胞的比例为10:1。将载玻片在高湿度下摇动(25 r.p.m.)1小时,然后用PBS冲洗三次,去除未附着的细菌。用台盼蓝(0.2%w/v28)淬灭非吞噬细菌的荧光。将带有细菌的血细胞在戊二醛-甲醛蒸汽中固定10分钟,并用PBS中的甘油(20%v/v)固定。必要时,血细胞被鉴定为颗粒细胞或浆细胞29。通过荧光显微镜测定总吞噬血细胞和浆细胞的水平(以总血细胞和总浆细胞的百分比表示)。使用相差显微镜测定每种附着细菌的血细胞类型的数量(附着包括吞噬细菌和血细胞表面的细菌)和每种血细胞类型附着细菌的数量。在包含10个样本的10个重复中,每个重复检查至少150个每种类型的血细胞。 血细胞蛋白排出量[1] 通过在冰上冷却10只幼虫(15分钟)并在1毫升冷冻抗凝剂中收集75µL血淋巴,获得不含血淋巴血浆的血细胞。通过离心洗涤血细胞三次,以去除血浆和抗凝剂(200g,2分钟)。然后将沉淀物重新悬浮在冷PBS(5°C,1 mL)中。将血细胞悬浮液(100µL)加入96孔微量滴定板的孔中,该微量滴定板含有圆形(直径5mm)玻璃盖玻片和PBS,含有和不含有选定浓度的Rp-8-Br-cAMPS和Sp-8-Br-cAMPS,或含有固定水平的激活剂和10和50 nmol/L的抑制剂。其他孔含有缓冲液和PKA调节剂,这两种细菌产生10:1的细菌:血细胞比。含有和不含酶调节剂的细菌作为无血细胞对照。将板在22°C下孵育15-60分钟,在此期间通过移液移取样品并离心(2000 g,2分钟)。测定上清液中的总蛋白。在整个培养过程中使用了10个重复,每个重复包含两个样本。 |
| 动物实验 |
体内PKA调节剂[1]
将包含两组各10只幼虫的5个重复组,在幼虫前胸足基部注射10 µL含有特定浓度Sp-8-Br-cAMPS和/或Rp-8-Br-cAMPS的PBS溶液。注射30分钟后,对其中一组幼虫进行采血,并使用血细胞计数器测定每只幼虫的总血细胞数。注射24小时后,对另一组幼虫进行解剖,以检测结节。同样,使用含有1 × 10⁸个细菌/10 µL的测试溶液,评估PKA调节剂对枯草芽孢杆菌和线虫黄单胞菌清除的影响。细菌和血细胞数量均使用血细胞计数器测定。 |
| 参考文献 | |
| 其他信息 |
我们单独使用蛋白激酶A (PKA) 特异性激活剂Sp-8-Br-cAMPS和I型抑制剂Rp-8-Br-cAMPS,以及二者联合使用,以明确PKA在体外和体内大蜡螟幼虫血细胞非自身反应中的作用。活性PKA抑制血细胞反应,而PKA抑制剂则增强多种反应,包括体外细菌吞噬作用、附着细菌的血细胞数量、细菌诱导的血细胞蛋白释放以及血细胞在载玻片上的黏附,以及体内血淋巴中细菌的清除。未黏附的血细胞比黏附的血细胞具有更高的PKA活性;因此,活性PKA限制了血细胞对异物的反应。我们发现:(i) 单独使用PKA抑制剂可诱导非自身反应,包括体内血细胞蛋白释放和血细胞数量减少,并诱导结节形成;(ii) 酶激活剂的作用与抑制剂的作用相反; (iii) 这些调节因子相互抵消,共同影响血细胞裂解液中的PKA活性。这些发现证实PKA是血细胞非自身反应的介质。[1] cAMP通过激活脂筏中组装的抑制性蛋白激酶A (PKA)-C端Src激酶通路,抑制导致T细胞活化的生化事件。在本研究中,我们证明Sp-8-溴-cAMPS(一种cAMP激动剂)激活I型PKA对周围效应T细胞中的抗原特异性免疫反应具有显著的抑制作用。I型PKA的激活以浓度依赖的方式抑制CD4+和CD8+ T细胞的细胞因子产生和增殖反应。观察到的cAMP效应似乎是内源性地发生在T细胞中,并且不依赖于抗原呈递细胞(APC)。免疫应答的抑制并非由特定T细胞凋亡引起,且可被PKA I型选择性cAMP拮抗剂逆转。这支持了PKA I型作为免疫应答负调控关键酶以及抑制自身反应性T细胞潜在靶点的观点。[2]我们和其他研究者已证实,cAMP和PKA I型在分子水平上可作为TCR/CD3信号通路的急性抑制剂。本研究的数据表明,cAMP通过激活PKA I型,成为T细胞功能的普遍抑制剂。Sp-8-Br-cAMPS预孵育可剂量依赖性地抑制抗原诱导的IFN-γ、TNF-α、IL-2和IL-4的表达。该抑制作用涉及CD4和CD8 T细胞,且不依赖于抗原呈递细胞(APC),也不会诱导细胞凋亡。此外,cAMP类似物抑制了SEB诱导的增殖以及对CMV和结核菌素PPD的抗原特异性免疫反应。与PKA I型选择性抑制剂Rp-8-Br-cAMPS共同孵育,可完全逆转PKA激动剂Sp-8-Br-cAMPS的抑制作用。这表明观察到的效应是由于PKA I型的激活所致,并且cAMP孵育后T细胞功能的抑制作用可及时被cAMP拮抗剂逆转。[2]
|
| 分子式 |
C10H11BRN5O5PS
|
|---|---|
| 分子量 |
424.17
|
| 精确质量 |
444.922
|
| CAS号 |
127634-20-2
|
| PubChem CID |
24757345
|
| 外观&性状 |
Typically exists as solid at room temperature
|
| LogP |
1.731
|
| tPSA |
182.5
|
| 氢键供体(HBD)数目 |
2
|
| 氢键受体(HBA)数目 |
10
|
| 可旋转键数目(RBC) |
1
|
| 重原子数目 |
24
|
| 分子复杂度/Complexity |
542
|
| 定义原子立体中心数目 |
4
|
| SMILES |
C1[C@@H]2[C@H]([C@H]([C@@H](O2)N3C4=NC=NC(=C4N=C3Br)N)O)OP(=S)(O1)[O-].[Na+]
|
| InChi Key |
SKJLJCVVXRNYGJ-QKAIHBBZSA-M
|
| InChi Code |
InChI=1S/C10H11BrN5O5PS.Na/c11-10-15-4-7(12)13-2-14-8(4)16(10)9-5(17)6-3(20-9)1-19-22(18,23)21-6;/h2-3,5-6,9,17H,1H2,(H,18,23)(H2,12,13,14);/q;+1/p-1/t3-,5-,6-,9-,22?;/m1./s1
|
| 化学名 |
sodium;(4aR,6R,7R,7aS)-6-(6-amino-8-bromopurin-9-yl)-2-oxido-2-sulfanylidene-4a,6,7,7a-tetrahydro-4H-furo[3,2-d][1,3,2]dioxaphosphinin-7-ol
|
| 别名 |
129735-00-8; 127634-20-2; 8-Bromoadenosine 3',5'-cyclic monophosphorothioate, Rp-isomer sodium salt; 8-Bromoadenosine 3',5'-cyclic monophosphorothioate, Sp-isomer sodium salt; 8-BROMOADENOSINE-3',5'-CYCLIC MONOPHOSPHOROTHIOATE, SP-ISOMER SODIUM SALT; Sp-8-Br-cAMPS*Na; 8-Br-cAMPS*Na Sp-isomer; YMC11590;
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
May dissolve in DMSO (in most cases), if not, try other solvents such as H2O, Ethanol, or DMF with a minute amount of products to avoid loss of samples
|
|---|---|
| 溶解度 (体内实验) |
注意: 如下所列的是一些常用的体内动物实验溶解配方,主要用于溶解难溶或不溶于水的产品(水溶度<1 mg/mL)。 建议您先取少量样品进行尝试,如该配方可行,再根据实验需求增加样品量。
注射用配方
注射用配方1: DMSO : Tween 80: Saline = 10 : 5 : 85 (如: 100 μL DMSO → 50 μL Tween 80 → 850 μL Saline)(IP/IV/IM/SC等) *生理盐水/Saline的制备:将0.9g氯化钠/NaCl溶解在100 mL ddH ₂ O中,得到澄清溶液。 注射用配方 2: DMSO : PEG300 :Tween 80 : Saline = 10 : 40 : 5 : 45 (如: 100 μL DMSO → 400 μL PEG300 → 50 μL Tween 80 → 450 μL Saline) 注射用配方 3: DMSO : Corn oil = 10 : 90 (如: 100 μL DMSO → 900 μL Corn oil) 示例: 以注射用配方 3 (DMSO : Corn oil = 10 : 90) 为例说明, 如果要配制 1 mL 2.5 mg/mL的工作液, 您可以取 100 μL 25 mg/mL 澄清的 DMSO 储备液,加到 900 μL Corn oil/玉米油中, 混合均匀。 View More
注射用配方 4: DMSO : 20% SBE-β-CD in Saline = 10 : 90 [如:100 μL DMSO → 900 μL (20% SBE-β-CD in Saline)] 口服配方
口服配方 1: 悬浮于0.5% CMC Na (羧甲基纤维素钠) 口服配方 2: 悬浮于0.5% Carboxymethyl cellulose (羧甲基纤维素) 示例: 以口服配方 1 (悬浮于 0.5% CMC Na)为例说明, 如果要配制 100 mL 2.5 mg/mL 的工作液, 您可以先取0.5g CMC Na并将其溶解于100mL ddH2O中,得到0.5%CMC-Na澄清溶液;然后将250 mg待测化合物加到100 mL前述 0.5%CMC Na溶液中,得到悬浮液。 View More
口服配方 3: 溶解于 PEG400 (聚乙二醇400) 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.3575 mL | 11.7877 mL | 23.5755 mL | |
| 5 mM | 0.4715 mL | 2.3575 mL | 4.7151 mL | |
| 10 mM | 0.2358 mL | 1.1788 mL | 2.3575 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
 Kiss2 peptide acetate
Kiss2 peptide acetate
 Rp-8-Br-cAMPS sodium
Rp-8-Br-cAMPS sodium
 8-Bromo-cAMP
8-Bromo-cAMP
 ISTH0036
ISTH0036
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 463611831
463611831
