| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| 500mg |
|
||
| 1g |
|
||
| Other Sizes |
|

| 靶点 |
NMDA receptor
|
|---|---|
| 体外研究 (In Vitro) |
在30分钟的固定应激或全身施用苯二氮卓/GABA(A)受体反向激动剂甲基-6,7-二甲氧基-4-乙基-β-卡博林-3-羧酸酯(DMCM)后,内侧前额叶皮层中的多巴胺代谢,如二羟基苯乙酸(DOPAC)浓度所反映的,显著增加。用苯二氮卓类/GABA(A)受体激动剂地西泮和唑吡坦预处理大鼠后,对应激的反应减弱。此外,用低效力部分激动剂R-(+)-3-氨基-1-羟基吡咯烷-2-酮(R-(+)-HA-966)和甘氨酸/NDA受体新型高亲和力全拮抗剂7-氯-4-羟基-3(3-苯氧基)苯基喹啉-2-(H)-酮(L-701324)预处理,可以减弱对应激和DMCM的反应。这些结果表明,甘氨酸/NDA受体复合物的拮抗剂在防止应激和GABA(A)受体反向激动剂激活大脑中皮层多巴胺系统的能力方面与苯二氮卓/GABA(A)接收器激动剂相当。讨论了甘氨酸/NDA受体拮抗剂、中脑皮层多巴胺系统和应激相关疾病之间的相互作用[3]。
|
| 体内研究 (In Vivo) |
在强迫游泳试验 (FST) 和悬尾试验 (TST) 中,L-701,324(5–10 mg/kg;腹腔注射;一次)显示出类似抗抑郁的潜力,且不影响小鼠的运动活动 [1]。在慢性不可预测的轻度应激 (CUMS) 抑郁模型中,L-701324(5-10 mg/kg;腹腔注射;每日一次,持续 2 周)显示出强烈的抗抑郁样作用,抑制 CUMS 诱导的欧洲生成,并减少 BDNF 信号级联反应。海马体[1]。 L-701324(2.5–5 mg/kg;口服;一次)减少无条件和无条件焦虑样行为,同时通过阻断 NMDA 受体上的 NMDA/甘氨酸敏感区域来抑制 NMDA 受体活性。包括有条件冲突行为的情况[2]。
目前临床上使用的抗抑郁药存在疗效低、起效慢、不良反应多等局限性。开发单胺能药物之外的新型抗抑郁药已成为必要。L-701,324是一种强效的NMDA受体拮抗剂,本研究旨在探讨L-701,324对小鼠可能的抗抑郁作用。在这里,我们同时使用了各种方法,包括强迫游泳试验(FST)、尾部悬吊试验(TST)、慢性不可预测的轻度应激(CUMS)抑郁模型、蛋白质印迹和免疫荧光。单次注射L-701324在FST和TST中表现出抗抑郁样潜力,而不影响小鼠的运动活动。反复注射L-701324不仅可以预防CUMS诱导的小鼠抑郁样行为,还可以改善CUMS对海马BDNF信号级联和神经发生的下调作用。此外,BDNF系统的强效抑制剂K252a完全阻断了L-701324在小鼠体内的抗抑郁样活性。K252a给药还消除了L-701324对CUMS治疗小鼠海马BDNF信号级联和神经发生的激活作用。总的来说,这些数据表明L-701324在小鼠体内具有抗抑郁样活性,这至少部分是通过促进海马BDNF系统介导的。[1] 在高架迷宫和Vogel冲突试验中检查了NMDA/甘氨酸位点拮抗剂7-氯-4-羟基-3-(3-苯氧基)苯基-2(1H)-喹诺酮(L-701324)和苯二氮卓类受体激动剂地西泮的作用。口服L-701324导致在开放臂中花费的时间百分比呈剂量依赖性增加(2.5和5.0 mg/kg,-30分钟),而进入+迷宫的臂总数或进入开放臂的百分比没有变化。相同剂量的L-701,324以剂量依赖的方式增加了Vogel冲突测试中的惩罚反应,对未受惩罚的饮酒行为没有影响。L-701324的抗焦虑样作用是在其本身对动物的运动活动没有影响的剂量下获得的。在正迷宫情况下,地西泮(2mg/kg,腹腔注射,-30分钟)比L-701324略有效,而在Vogel测试中,两种化合物的惩罚饮酒增加幅度相同。我们目前的研究结果表明,通过阻断NMDA受体上的NMDA/甘氨酸敏感位点来抑制NMDA受体活性,在非条件性和条件性冲突行为情境中都伴随着焦虑样行为的减少。[2] 甘氨酸/NDA受体拮抗剂R-(+)-HA-966和L-701324对应激诱导的中脑DOPAC浓度增加的影响[3] 对大鼠进行R-(+)-HA-966(20mg/kg,i.p.)或L-701,324(5mg/kg,i.p..)预处理,可显著减轻30分钟固定应激后内侧前额叶皮层DOPAC浓度的增加,而不会影响DOPAC浓度本身(图3a-c)。相比之下,用L-701357(10mg/kg,i.p.)预处理,L-701324的非活性对映体,对内侧前额叶皮层中DOPAC浓度的基础或应激诱导的增加没有显著影响(图3d)。内侧前额叶皮层中的多巴胺浓度不受压力或药物治疗的影响(数据未显示)。 甘氨酸/NDA受体拮抗剂L-701,324对苯二氮卓/GABA受体反向激动剂DMCM诱导的中脑DOPAC浓度增加的影响[3] 在给予GABAA受体反向激动剂DMCM(5mg/kg,i.p.)后,用L-701,324(5mg/kg)对大鼠进行预处理显著减轻了内侧前额叶皮层DOPAC浓度的增加。相比之下,用L-701357(10mg/kg,i.p.)预处理,L-701324的非活性区域异构体,不影响DMCM增加内侧前额叶皮层DOPAC浓度(图4)。内侧前额叶皮层中的多巴胺浓度不受DMCM或L-701324的影响(数据未显示)。 |
| 动物实验 |
动物/疾病模型: 雄性 C57BL/6 J 小鼠(7 周龄),慢性不可预测轻度应激 (CUMS) [1]
剂量: 5 和 10 mg/kg 给药途径: 腹腔注射 (ip);每日一次,连续 2 周 实验结果: C57BL/6 J 小鼠的静止不动时间减少。海马中 BDNF、pTrkB 和 pCREB 的表达增加。 动物/疾病模型: 雄性 C57BL/6 J 小鼠,强迫游泳试验 (FST) 和悬尾试验 (TST)(7 周龄)[1] 剂量: 5 和 10 mg/kg 给药途径: 腹腔注射 (ip); 实验结果: C57BL/6J 小鼠在强迫游泳试验 (FST) 和悬尾试验 (TST) 中的不动时间减少。 动物/疾病模型: 雄性 SD(Sprague-Dawley)大鼠(280-300 g)[2] 剂量: 2.5 和 5 mg/kg 给药途径: 口服; 给药途径: 口服。 实验结果: 小鼠张开手臂的时间百分比呈剂量依赖性增加。在 Vogel 冲突测试中,惩罚反应呈剂量依赖性增加。 L-701,324 溶于含 1% DMSO(溶剂)的生理盐水中,并腹腔注射 (ip) (10 mL/kg)。 L-701,324(5 和 10 mg/kg)、氟西汀(20 mg/kg)和 K252a(25 μg/kg)的剂量根据之前的报道和我们的预实验[1]确定。强迫游泳试验 (FST) [1] 在试验前 30 分钟,对未经处理的 C57BL/6J 小鼠进行单次腹腔注射 L-701,324、氟西汀或溶剂。将小鼠单独放入一个透明玻璃缸(内含 15 cm 高的纯水,温度为 25 ± 1 °C)中 6 分钟。使用秒表记录每只小鼠在最后 4 分钟内的静止时间。每次试验后更换水。静止的定义为小鼠漂浮在水中,没有挣扎或仅有轻微的动作以保持鼻子露出水面。这项测试是在观察者不知晓实验分组的情况下进行的。 悬尾测试 (TST) [1] 在测试前 30 分钟,对未经处理的 C57BL/6 J 小鼠进行单次腹腔注射 L-701,324、氟西汀或载体。将每只小鼠的尾尖分别粘在离地 60 厘米的横杆上,并悬挂 6 分钟。记录每只小鼠在这 6 分钟内的静止(完全不动)时间。这项测试是在观察者不知晓实验分组的情况下进行的。 旷场测试 (OFT) [1] 这项测试在暗室中进行。在测试前 30 分钟,对未经处理的 C57BL/6 J 小鼠进行单次腹腔注射 L-701,324、氟西汀或载体。将小鼠单独放置在开放式场地装置(100 × 100 × 45 cm;25 个方格,每个方格 20 × 20 cm)的地面上,允许其自由探索 5 分钟。装置顶部装有红色灯泡(50 W)照明。记录每只小鼠在 5 分钟内穿过的方格数。该测试的记录者不知晓小鼠的实验分组。每次测试后,清洁地面。 慢性不可预测轻度应激 (CUMS) [1] 简而言之,本研究采用了 8 种应激源:潮湿垫料(24 小时)、笼子倾斜(12 小时)、束缚(1 小时)、摇晃(30 分钟)、4 °C 暴露(1 小时)、昼夜颠倒、食物剥夺(23 小时)或饮水剥夺(23 小时)。所有这些应激源均随机给予,持续6周,并在最后2周每日给予L-701,324/氟西汀/载体。对照组小鼠除常规操作(例如定期清洁笼子)和药物处理外,不进行其他干扰。在慢性不可预测性应激(CUMS)后,同时进行强迫游泳试验(FST)、悬尾试验(TST)和蔗糖偏好试验,以评估动物的抑郁样行为。 高架十字迷宫实验[2] 十字形迷宫由木材制成,放置在安静的实验室环境中,距离地面50厘米。迷宫的两个相对臂是开放的(50×10厘米),另外两个臂被墙壁封闭(50×10×40厘米)。实验在黑暗安静的房间内进行,房间上方80厘米处有一盏15瓦的恒定光源照射迷宫装置。开放臂和封闭臂的光照强度相同。实验前三天,每天对每只大鼠进行5分钟的抓取操作。实验前60分钟,将动物连同笼子一起放入单独的安静房间。在开始十字迷宫行为记录之前,将每只动物放入一个新环境中(以传统的斯金纳箱为代表),持续5分钟。十字迷宫实验开始时,将大鼠放入十字迷宫的中心,使其面向一个开放臂。随后,由一位对药物处理方案不知情的独立观察员记录大鼠进入两个臂的次数和停留时间,持续5分钟。当大鼠四肢全部进入臂内时,记录一次“进入臂”。每次测试结束后,用自来水仔细清洗迷宫;当天所有实验结束后,用稀释的酒精溶液清洗迷宫。开放臂的活动量化指标包括:a) 在开放臂停留的时间;b) 进入开放臂的次数。甘氨酸受体拮抗剂L-701,324在测试前30分钟经口(PO)给予。 Vogel冲突测试[2] 饮水训练和冲突抑制饮水实验(实验方案见Möller等人,1997)在两个标准操作行为箱中进行,但这些箱子是专门为电击诱导大鼠饮水抑制而设计的。整个实验周期为3天。第一天早上,将动物笼中的饮用水移除。第二天和第三天,将受试动物放入测试装置中,允许其自由饮水12分钟,期间不施加电击。经过这两天的训练后,大多数动物在训练期间记录的舔舐次数达到稳定的基线水平。少数动物拒绝饮水,因此被移出实验。实验当天,将动物随机分为对照组(注射溶剂)和实验组(注射目标药物)。给药30分钟后,将动物放入实验装置中,待动物舔舐水瓶20次后,实验正式开始。此后,若动物继续饮水,则会受到电击,电击以5秒为周期,间隔4秒,电流强度为0.2 mA。记录12分钟内受惩罚和未受惩罚的饮水次数,并将受惩罚的饮水次数作为抑制饮水行为的指标。所有记录均在凌晨12点至下午6点之间进行,以避免昼夜节律对结果造成较大影响。苯二氮卓受体激动剂地西泮以2.0 mg/kg的剂量腹腔注射(IP)给药,该剂量此前已被证实可有效抑制大鼠的情绪。在行为记录开始前 30 分钟,以 2.5 和 5.0 mg/kg 的剂量口服给予甘氨酸受体拮抗剂 L-701,324。 L-701,324 以 0.5% 甲基纤维素溶液配制的悬浮液形式口服。 L-701,324(1 和 5 mg/kg,腹腔注射)、L-701,357(10 mg/kg,腹腔注射)或载体(0.5% 羧甲基纤维素溶于 0.9% 生理盐水,1 ml/kg,腹腔注射)。 30分钟后,大鼠分别腹腔注射DMCM(5 mg/kg)或溶剂(1 ml/kg),30分钟后处死;或在应激实验中,大鼠被留在笼内或固定30分钟后立即处死。取出脑组织,分离内侧前额叶皮层,用固态二氧化碳冷冻,并储存于-70℃。所有脑组织样本均采用高效液相色谱-电化学检测法(HPLC-ECD)分析多巴胺及其酸性代谢物二羟基苯乙酸(DOPAC)的含量(Hutson等,1991)。简而言之,将组织样本在10倍体积的0.4 M高氯酸溶液中匀浆,该溶液含有0.1%半胱氨酸、0.01%焦亚硫酸钠和0.01%乙二胺四乙酸钠(NaEDTA),并在3000×g下离心10分钟。高效液相色谱系统采用 HPLC Technology Techsphere 3μ ODS 色谱柱(4.6 mm×7.5 cm)。流动相由 0.07 M KH2PO4、0.0035% NaEDTA、0.023% 辛基硫酸钠和 12.5% 甲醇组成,pH 值为 2.75,流速为 1 ml/min。多巴胺及其代谢物采用 Antec 电化学检测器(Presearch)进行检测,工作电极相对于银/氯化银参比电极的电位设置为 +0.65 V。[3] |
| 参考文献 |
|
| 其他信息 |
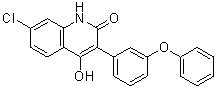
7-氯-4-羟基-3-(3-苯氧基苯基)-1H-喹啉-2-酮属于喹啉类化合物。
内侧前额叶皮层中多巴胺代谢(以二羟基苯乙酸 (DOPAC) 的浓度反映)在30分钟固定应激或全身性给予苯二氮卓类/GABA(A)受体反向激动剂甲基-6,7-二甲氧基-4-乙基-β-咔啉-3-羧酸酯 (DMCM) 后显著增加。预先用苯二氮卓类/GABA(A)受体激动剂地西泮和唑吡坦处理大鼠可减弱应激反应。此外,用低效部分激动剂R-(+)-3-氨基-1-羟基吡咯烷-2-酮(R-(+)-HA-966)和新型高亲和力甘氨酸/NMDA受体完全拮抗剂7-氯-4-羟基-3-(3-苯氧基)苯基喹啉-2-(H)-酮(L-701,324)进行预处理,可减弱应激和DMCM引起的反应。这些结果表明,甘氨酸/NMDA受体复合物拮抗剂在抑制应激和GABA(A)受体反向激动剂激活中脑皮质多巴胺系统方面,与苯二氮卓类/GABA(A)受体激动剂具有可比性。研究结果探讨了甘氨酸/NMDA受体拮抗剂、中脑皮层边缘多巴胺系统以及应激相关疾病之间的相互作用。[3] L-701,324治疗促进海马神经发生的发现令人振奋,表明L-701,324可能是一种促神经发生化合物。鉴于BDNF与神经发生之间存在明确的相关性,L-701,324很可能通过增强海马BDNF的表达来调节神经发生。L-701,324也可能调节一些已知的促神经发生因子,例如性别决定区Y盒2(SDR2)和配对盒蛋白6(P6),这需要进一步研究。抑郁症的神经生物学机制相当复杂。除了脑源性神经营养因子(BDNF)功能障碍、单胺类神经递质功能减退和下丘脑-垂体-肾上腺轴(HPA轴)功能亢进外,近期研究越来越多地报道了其他与抑郁症相关的靶点,例如过氧化物酶体增殖激活受体α(PPARα)、血管内皮生长因子(VEGF)、盐诱导激酶2(SIC2)和ΔFosB。尽管涉及K252a的实验结果表明BDNF参与了L-701,324的抗抑郁样作用,但我们不能排除除BDNF以外的其他蛋白靶点。总之,L-701,324在小鼠中具有抗抑郁样作用,其作用机制至少部分是通过增强海马BDNF信号通路实现的。 [1] 本研究发现,甘氨酸/NMDA 受体的完全拮抗剂L-701,324可减弱应激和苯二氮卓类/GABAA 受体反向激动剂引起的皮质多巴胺代谢变化,这与谷氨酸,特别是 NMDA 受体复合物参与应激和焦虑反应的证据相一致。因此,行为学研究表明,NMDA受体复合物谷氨酸、甘氨酸和离子通道位点的拮抗剂在啮齿动物的条件反射和非条件反射测试中均表现出抗焦虑活性(Trullas et al., 1989; Corbett and Dunn, 1991; Dunn et al., 1992; Kehne et al., 1991; Faiman et al., 1994)。本研究的结果也支持先前的发现,即NMDA受体对士的宁不敏感的甘氨酸位点的拮抗剂与中脑皮层边缘多巴胺神经元相互作用,但仅在这些神经元被激活时才起作用。因此,研究表明,R-(+)-HA-966 和 L-701,324 不仅不具备非竞争性离子通道阻滞剂苯环利定和 MK801 增加中脑皮层边缘多巴胺代谢和诱导活动过度的能力,而且还能显著减弱 PCP 和 MK801 的这些作用(Bristow 等,1993;Bristow 等,1996a;Hutson 等,1991;Hutson 等,1995)。基于这些研究和其他研究,有研究提示甘氨酸/NMDA 受体拮抗剂在啮齿动物中表现出非典型的抗精神病药物样特性。鉴于额叶皮层在精神分裂症的病理生理学中的重要性,以及压力可能是精神分裂症患者复发(Dohrenwend 和 Egri,1981)和精神分裂症症状加重(Bebbington 等,1993)的缓解因素,本研究结果强调了先前的观点,即谷氨酸系统可能在精神分裂症中发挥重要作用,并且这类化合物可能是一种治疗精神分裂症的新方法。此类化合物在治疗精神分裂症或焦虑相关疾病方面是否具有临床疗效,仍有待确定。[3] |
| 分子式 |
C21H14CLNO3
|
|---|---|
| 分子量 |
363.7938
|
| 精确质量 |
363.066
|
| 元素分析 |
C, 69.33; H, 3.88; Cl, 9.75; N, 3.85; O, 13.19
|
| CAS号 |
142326-59-8
|
| PubChem CID |
54682505
|
| 外观&性状 |
Typically exists as White to off-white solids at room temperature
|
| 密度 |
1.4±0.1 g/cm3
|
| 沸点 |
584.7±50.0 °C at 760 mmHg
|
| 闪点 |
307.4±30.1 °C
|
| 蒸汽压 |
0.0±1.7 mmHg at 25°C
|
| 折射率 |
1.680
|
| LogP |
5.5
|
| tPSA |
62.32
|
| 氢键供体(HBD)数目 |
2
|
| 氢键受体(HBA)数目 |
3
|
| 可旋转键数目(RBC) |
3
|
| 重原子数目 |
26
|
| 分子复杂度/Complexity |
558
|
| 定义原子立体中心数目 |
0
|
| SMILES |
ClC1C([H])=C([H])C2C(=C(C(N([H])C=2C=1[H])=O)C1C([H])=C([H])C([H])=C(C=1[H])OC1C([H])=C([H])C([H])=C([H])C=1[H])O[H]
|
| InChi Key |
FLVRDMUHUXVRET-UHFFFAOYSA-N
|
| InChi Code |
InChI=1S/C21H14ClNO3/c22-14-9-10-17-18(12-14)23-21(25)19(20(17)24)13-5-4-8-16(11-13)26-15-6-2-1-3-7-15/h1-12H,(H2,23,24,25)
|
| 化学名 |
7-chloro-4-hydroxy-3-(3-phenoxyphenyl)quinolin-2(1H)-one
|
| 别名 |
L701,324; L701324; L 701324; L 701,324; 142326-59-8; L-701,324; L-701,324; 7-chloro-4-hydroxy-3-(3-phenoxyphenyl)quinolin-2(1H)-one; CHEMBL31741; 2(1H)-Quinolinone, 7-chloro-4-hydroxy-3-(3-phenoxyphenyl)-; 7-CHLORO-4-HYDROXY-3-(3-PHENOXYPHENYL)-1H-QUINOLIN-2-ONE; I9WY146163; L-701324; L-701,324.
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
DMSO : ≥ 34 mg/mL (~93.46 mM)
|
|---|---|
| 溶解度 (体内实验) |
注意: 如下所列的是一些常用的体内动物实验溶解配方,主要用于溶解难溶或不溶于水的产品(水溶度<1 mg/mL)。 建议您先取少量样品进行尝试,如该配方可行,再根据实验需求增加样品量。
注射用配方
注射用配方1: DMSO : Tween 80: Saline = 10 : 5 : 85 (如: 100 μL DMSO → 50 μL Tween 80 → 850 μL Saline)(IP/IV/IM/SC等) *生理盐水/Saline的制备:将0.9g氯化钠/NaCl溶解在100 mL ddH ₂ O中,得到澄清溶液。 注射用配方 2: DMSO : PEG300 :Tween 80 : Saline = 10 : 40 : 5 : 45 (如: 100 μL DMSO → 400 μL PEG300 → 50 μL Tween 80 → 450 μL Saline) 注射用配方 3: DMSO : Corn oil = 10 : 90 (如: 100 μL DMSO → 900 μL Corn oil) 示例: 以注射用配方 3 (DMSO : Corn oil = 10 : 90) 为例说明, 如果要配制 1 mL 2.5 mg/mL的工作液, 您可以取 100 μL 25 mg/mL 澄清的 DMSO 储备液,加到 900 μL Corn oil/玉米油中, 混合均匀。 View More
注射用配方 4: DMSO : 20% SBE-β-CD in Saline = 10 : 90 [如:100 μL DMSO → 900 μL (20% SBE-β-CD in Saline)] 口服配方
口服配方 1: 悬浮于0.5% CMC Na (羧甲基纤维素钠) 口服配方 2: 悬浮于0.5% Carboxymethyl cellulose (羧甲基纤维素) 示例: 以口服配方 1 (悬浮于 0.5% CMC Na)为例说明, 如果要配制 100 mL 2.5 mg/mL 的工作液, 您可以先取0.5g CMC Na并将其溶解于100mL ddH2O中,得到0.5%CMC-Na澄清溶液;然后将250 mg待测化合物加到100 mL前述 0.5%CMC Na溶液中,得到悬浮液。 View More
口服配方 3: 溶解于 PEG400 (聚乙二醇400) 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.7488 mL | 13.7442 mL | 27.4884 mL | |
| 5 mM | 0.5498 mL | 2.7488 mL | 5.4977 mL | |
| 10 mM | 0.2749 mL | 1.3744 mL | 2.7488 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
 NMDAR antagonist 3
NMDAR antagonist 3
 Pep1-AGL
Pep1-AGL
 NMDA receptor modulator 9
NMDA receptor modulator 9
 Benzylaspartic acid
Benzylaspartic acid
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1































 COA
COA
 463611831
463611831
