| 规格 | 价格 | |
|---|---|---|
| 500mg | ||
| 1g | ||
| Other Sizes |
| 靶点 |
Arachidonic acid metabolite
|
|---|---|
| 体外研究 (In Vitro) |
12-HETE激活 ILK/NF-κB 通路,有助于抑制细胞凋亡,并提示维持卵巢癌细胞活力的关键潜在机制。 12-HETE 通过触发整合素连接激酶/NF-κB 通路促进卵巢癌细胞存活。 12-HETE 对卵巢癌细胞凋亡具有浓度依赖性保护作用。 12-HETE (1 µM) 显着降低血清剥夺 (SD) 诱导的 caspase-3 激活。 12-HETE 的 IC50 值为 1.13 µM,以浓度依赖性方式抑制 SD 产生的 caspase-3 活性增加[1]。在卵巢癌细胞中,12-HETE (1 µM) 通过 ILK 促进 NF-κB 激活和核转位 [1]。 12-HETE 会导致人类胰岛细胞死亡,降低代谢活性,并减少胰岛素分泌。 12-HETE 抑制前列腺素 E1 诱导的细胞内 cAMP 水平升高,同时增强凝血酶诱导的牛血小板聚集。 12-HETE 可防止洗涤后的血小板 (WP) 聚集[2]。 12-HETE 对神经元有多种影响,包括减少谷氨酸释放和钙流入以及抑制 AMPA 受体 (AMPA-R) 激活[3]。
12-HETE对细胞凋亡的影响是通过ILK途径介导的[1] 我们之前的研究表明,ILK参与调节卵巢癌症的进展,并在癌症细胞的存活中发挥关键作用。15我们研究了12-HETE是否在卵巢癌症的进展过程中负责ILK途径的激活。在OVCAR-3细胞中,用1µM 12-HETE处理导致ILK表达增加(图2A)。为了阐明ILK在12-HETE抑制细胞凋亡中的作用,使用ILK siRNA(siILK)敲低ILK的表达(图2B)。如图S1所示,与未转染的OVCAR-3细胞相比,siControl转染对细胞存活率、半胱氨酸天冬氨酸蛋白酶-3活性和ILK表达没有明显影响(图S1A-C),这意味着siControl转染和未转染细胞之间没有显著差异。因此,我们在以下实验中使用siControl转染细胞作为阴性对照。细胞生长试验的结果表明,1µM 12-HETE拮抗了SD诱导的细胞存活率的降低,但在用siILK抑制ILK表达后,12-HETE对细胞存活的影响被消除了(图2C)。如图2D所示,1µM 12-HETE减弱了SD诱导的LDH释放(细胞死亡的指示),而ILK siRNA消除了12-HETE在LDH释放中的作用(图2D)。此外,ILK siRNA减弱了12-HETE对SD诱导的半胱氨酸天冬氨酸蛋白酶-3激活的抑制作用和切割半胱氨酸天冬氨酰蛋白酶-3表达的增加(图2E和F)。这些结果表明12-HETE通过激活ILK途径促进卵巢癌症细胞的存活。 ILK途径的12-HETE激活抑制线粒体依赖性凋亡[1] 固有细胞凋亡是SD诱导的导致细胞生长停滞的重要事件。我们研究了线粒体依赖性凋亡的一些重要变化,包括Bcl-2和Bax的蛋白水平、细胞色素C释放到细胞质中以及胱天蛋白酶-9的活性。如图3A和B所示,1µM 12-HETE诱导Bcl-2(一种抗凋亡蛋白)的表达并抑制Bax(一种促凋亡蛋白),这些作用被ILK siRNA削弱。此外,细胞色素C从线粒体释放到细胞质中是触发线粒体依赖性凋亡进展的关键事件。我们发现SD处理导致细胞质细胞色素C的表达增加,线粒体细胞色素C表达减少,但1µM 12-HETE减轻了细胞色素C从线粒体释放到细胞质中的过程。然而,ILK siRNA消除了12-HETE对细胞色素C释放的抑制作用(图3C)。此外,我们还检测了胱天蛋白酶-9的活性,胱天蛋白酶9在内在凋亡途径中被特异性激活。我们发现1µM 12-HETE抑制了SD诱导的胱天蛋白酶-9的激活,而ILK siRNA削弱了12-HETE对胱天蛋白酶-9-激活的抑制作用(图3D)。这些结果表明,ILK参与了12-HETE对线粒体依赖性凋亡的抑制。 同时,为了验证12-HETE在细胞凋亡中的作用,我们还尝试了额外浓度的12-HETE用于一些功能实验。我们的结果表明,用3µM 12-HETE处理可诱导OVCAR-3细胞中ILK的表达(图S2A)。3µM 12-HETE可以缓解SD诱导的细胞存活率降低和LDH释放增加。然而,ILK siRNA拮抗了12-HETE在细胞存活中的促进作用(图S2B和C)。此外,用3µM 12-HETE处理抑制了SD诱导的半胱氨酸天冬氨酸蛋白酶-3的激活,并增加了Bax的表达,但这些作用被ILK siRNA消除了(图S2D和E)。 通过沉默ILK的表达消除12-HETE诱导的NF-κB的激活[1] NF-κB是一种普遍存在的转录因子,其功能障碍与不同类型的癌症密切相关。NF-κB的激活增加是疾病进展不良的预测因素,并赋予细胞凋亡抵抗力。16基于这一证据,我们检测了12-HETE治疗条件下卵巢癌症细胞中NF-κB的状态。结果表明,1µM 12-HETE导致NF-κB p65磷酸化水平升高,并且12-HETE引起的NF-κBp65的磷酸化增加被ILK的敲除所减弱(图4A)。此外,细胞质和核提取物中NF-κB p65的蛋白水平表明,1µM 12-HETE处理导致核NF-κBp65蛋白水平显著升高,同时细胞质中NF-εBp65水平降低。然而,ILK siRNA消除了12-HETE治疗引起的核转位(图4B)。此外,NF-κB的激活受到IKB激酶的密切调节。因此,我们进一步研究了12-HETE对IKBa蛋白磷酸化的影响。如图4C所示,1µM 12-HETE促进了IKBa的磷酸化,这被ILK siRNA减弱。这些结果表明,12-HETE通过ILK促进了卵巢癌症细胞中NF-κB的激活和核易位。 此外,为了避免ILK siRNA的非特异性抑制,我们使用了ILK的另一种独立siRNA(siILK#2),并重复了一些重要的实验。Western blot证实了siILK#2的敲除效率(图S3A)。我们发现1µM 12-HETE对SD诱导的细胞存活率降低和半胱氨酸天冬氨酸蛋白酶-3活性增加的抑制作用被siILK#2缓解(图S3B和C)。此外,12-HETE抑制Bax的表达并增加NF-κB p65的磷酸化,但在用siILK#2抑制ILK的表达后,这些作用被消除了(图S3D和E)。 12-HETE对细胞凋亡的抑制作用被NF-κB的抑制所削弱[1] 为了说明NF-κB在12-HETE抑制细胞凋亡中的关键作用,我们使用5µM Bay-117082阻断卵巢癌症细胞中的NF-κB途径。17如图5A所示,1µM 12-HETE引起的细胞活力增加被NF-κB-抑制剂拮抗。此外,12-HETE对SD诱导的胱天蛋白酶-3和胱天蛋白酶-9激活的抑制作用因NF-κB的抑制而减弱(图5B和C)。此外,Bay-117082缓解了12-HETE介导的Bax蛋白水平的降低(图5D)。这些结果表明12-HETE通过ILK/NF-κB途径在卵巢癌症中保护细胞不凋亡。 12-HETE通过ILK和NF-κB抑制SKOV3卵巢癌症细胞的细胞生长和凋亡[1] 为了确定12-HETE的抗凋亡作用是否对OVCAR-3细胞具有特异性,我们检测了其在SKOV3卵巢癌症细胞中的作用。我们的结果表明,1µM 12-HETE抑制了SD诱导的细胞存活率的降低,并且这种作用被siILK减弱(图6A)。1µM 12-HETE处理可抑制SKOV3细胞中半胱氨酸天冬氨酸蛋白酶-3的激活和Bax蛋白水平的升高,而12-HETE对细胞凋亡的抑制作用可通过敲除ILK来减轻(图6B和C)。此外,1µM 12-HETE抑制了细胞存活率的降低,12-HETE介导的半胱氨酸天冬氨酸蛋白酶-3活性和Bax表达的增加受到NF-κB途径抑制剂的阻碍(图6D-F)。所有上述结果表明12-HETE通过调节卵巢癌症细胞中的ILK/NF-κB途径来促进细胞存活。 在AMPA-R介导的毒性模型中,钙可以通过AMPA-R和VSCC进入神经元(Colwell和Levine,1999;Leski等人,1999)。为了验证12-HETE的保护作用是否由这种去极化敏感通道介导,我们在250 nm的钙奎定存在下进行了毒性研究。钙奎定是一种能够灭活L-、N-和P-型钙通道的毒素(Schweitz等人,1994)。钙奎醚治疗降低了谷氨酸刺激后释放的LDH的总体水平(图6A),但也完全阻止了12-(S)HETE的保护作用,这表明12-HETE和钙奎醚可能共享一个作用位点。为了证实这一结果并进一步表征对12-HETE抑制敏感的钙通道群体,用通道特异性抑制剂重复了这项研究。在N型阻断剂芋螺毒素GVIa(500 nm,图6B)或合成L型通道阻断剂硝苯地平(2μm,图6C)存在的情况下,所取得的结果与用卡奎定观察到的结果相似(即VSCC阻断剂降低了毒性,而12-HETE没有提供额外的保护)。相比之下,尽管P/Q型钙通道毒素agatoxin IVa(250 nm)降低了谷氨酸的毒性,但其保护作用与12-HETE的保护作用是相加的(图6D),表明agatoxin和12-HETE影响不同的靶点。这些数据强烈表明,12-HETE通过抑制N型和L型VSCC来预防谷氨酸诱导的兴奋毒性,但不影响P型通道功能。 |
| 体内研究 (In Vivo) |
12-HETE也被证明可以增加凝血酶诱导的牛血小板聚集,这种作用抑制了前列腺素E1诱导的细胞内cAMP水平的升高。12-HETE增加凝血酶诱导的血小板聚集的能力在人类受试者中得到证实,这与[Ca2+]动员有关。为了更好地理解凝血酶对12-HETE释放的影响,Holinstat及其同事表征并比较了凝血酶受体、蛋白酶激活受体(标准杆数)1和4的激活对PLA2活性的影响。作者表明,在血小板中,两种不同的PAR诱导的信号通路为COX或LOX酶提供AA。这些作者探讨了凝血酶或PAR1和4激活肽(AP)激活WP后12-HETE形成的动力学:与TXB2相比,12-HETE的形成明显延迟。特别是,PAR1-AP诱导12-HETE的立即形成,而这种二十烷类化合物的形成在血小板暴露于PAR4-AP后延迟。此外,虽然PAR1诱导的TXB2形成在15秒内达到最大水平,但12-HETE生物合成持续的时间更长,仅在120秒内达到平台。这些数据可以用COX-1是自杀酶的概念来想象,而12-LOX随着时间的推移继续氧化AA,其活性可被视为底物AA可用性的函数。
Burzaco及其同事最近进行的一项研究也证实了凝血酶增加12-HETE水平的能力,该研究进一步证明了凝血酶增加人类WP中12-HETE生物合成的能力。在这项研究中,细胞外ATP以剂量依赖的方式抑制凝血酶诱导的血小板聚集,并抑制肌醇磷酸盐的产生和细胞内[Ca2+]的动员。 关于胶原蛋白诱导的血小板活化,Sekiya表明,添加到牛血小板中的外源性12-HETE抑制了磷脂中AA的释放和胶原蛋白诱导的抗血小板聚集,这与Takenaga之前的研究一致[2]。 |
| 酶活实验 |
单个神经元胞浆钙浓度([Ca2+]i)的测量。[3]
神经元以1×106/cm2的密度接种在玻璃盖玻片上。培养8天后,在HEPES缓冲的克雷布斯-林格盐水(KRS)中洗涤细胞两次,该盐水含有(单位为毫米):125 NaCl、5 KCl、1 Na2HPO4、1 MgSO4、1 CaCl2、5.5葡萄糖和20 HEPES,pH 7.2。然后将盖玻片置于4μm的fura-2AM KRS溶液中,在室温下持续轻轻搅拌22分钟。接下来,洗涤细胞并在新鲜KRS中再孵育22分钟(Grimaldi和Cavallaro,1999)。然后将盖玻片安装在一个腔室中,以约800μl/min的流速用含有0.002%甲基纤维素(载体)的KRS灌注。通过配备40×neofluar透镜和连接到台式计算机的增强CCD相机的倒置显微镜采集图像。Metafluor软件(宾夕法尼亚州西切斯特通用成像公司)用于分析实验数据。通过交替激发波长为340和380nm的制剂并记录波长为510nm的发射光获得的比值,使用Grynkiewicz方程(减法方程形式)转换为钙浓度(Grynkiewicz等人,1985)。该方程的校准值(Rmax−R,R−Rmin)是通过分别将神经元暴露于含有10 mm钙的KRS中的10μ米诺霉素或无钙KRS中20 mm EGTA中获得的。 细胞内cAMP测量。[3] 使用市售酶免疫测定试剂盒测量细胞内cAMP水平。在测定之前,将皮质神经元培养物暴露于100μm异丁基甲基黄嘌呤和不同浓度的12-HETE中10分钟,并在暴露于5μm毛喉素期间暴露15分钟。在此之后,用测定制造商提供的洗涤剂溶液裂解细胞,并评估细胞内cAMP浓度。根据制造商的说明进行了检测。 激动剂刺激的[35S]GTPγS结合研究。[3] 在含有0.32米蔗糖、50毫米Tris-HCl、pH 7.4、3毫米MgCl2和1毫米EGTA(pH 7.4)的缓冲液中,在冰上破坏大鼠前脑(不包括小脑)。将匀浆在1000×g下离心10分钟,丢弃颗粒,然后将上清液在3000×g下重新离心30分钟。将所得颗粒在冰冷的Tris缓冲液中研磨,该缓冲液含有(以毫米计):50 Tris-HCl,pH 7.4,3 MgCl2和1 EGTA,pH 7.4;然后在3000×g下重新离心30分钟。然后将颗粒悬浮在10体积的含有0.03%洋地黄皂苷的Tris缓冲液中,在室温下孵育15分钟,在3000×g下离心30分钟,然后将膜分散在Tris缓冲溶液中;使用Bradford试剂测定蛋白质浓度,然后将膜在-80°C下冷冻成等分。在测定当天,将等分试样解冻并稀释在含有(以毫米计):20 HEPES、100 NaCl、3 MgCl2和0.5 EGTA以及0.1%(w/v)BSA(pH 7.4)的测定缓冲液中。然后将等分试样在3000×g下离心30分钟。将膜亚液化到1毫升含有15μg蛋白质、30μm GDP、160 pm[35S]GTPγS和不同浓度12-(S)HETE/12-HETE的测定缓冲液中。在30μm“冷”GTPγS存在下测定非特异性结合。样品在23°C下孵育1小时,之后通过Whatman GF/B玻璃纤维过滤器(Whatman,Maidstone,UK)在真空下快速过滤终止反应,然后用冷测定缓冲液洗涤四次。通过液体闪烁计数测定与过滤器结合的放射性。绘制数据并使用Prism软件计算EC50值。 |
| 细胞实验 |
细胞活力测定[1]
细胞类型:卵巢癌 OVCAR-3 和 SKOV3 细胞 测试浓度: 0、0.2、0.5 和 1 µM 孵育时间:0、24、48、72 和 96 小时 实验结果:抑制 SD 诱导的细胞活力下降剂量依赖性方式。 1 µM 12-HETE处理显着减轻了 SD 条件下细胞活力的下降。 蛋白质印迹分析[1] 细胞类型:卵巢癌 OVCAR-3 和 SKOV3 细胞 测试浓度: 1 µM 孵育持续时间: 实验结果:导致 NF-κB p65 磷酸化水平增加。导致核 NF-κB p65 蛋白水平显着增加,同时细胞质中 NF-κB p65 水平降低。 MTT法[1] 细胞(OVCAR-3和SKOV3)在96孔板中以每孔约1×104个细胞的密度培养。在10%FBS DMEM培养基中培养8-12小时后,将细胞在无血清的DMEM中培养过夜,并在无FBS的低血糖DMEM中按组用指定试剂处理(血清剥夺[SD])。每24小时以指定浓度加入溶剂控制剂和其他试剂。将细胞作为不同组处理48小时后,在37°C下在0.5%MTT中孵育4小时,MTT是一种黄色线粒体染料,溶解在无菌PBS缓冲液中,然后通过将细胞与DMSO孵育10分钟来终止反应。测量540nm处的分光光度计吸光度。MTT形成的蓝色甲赞染料的量与存活细胞的数量成正比。 LDH测定[1] LDH的活性用于检测释放到培养基中的LDH水平,并通过Beyotime生物技术研究所的细胞毒性检测试剂盒进行测量。通过将培养基的LDH活性与完全细胞裂解后的LDH活性或总LDH活性进行比较,确定培养物中受损细胞的比例。使用含有Triton裂解细胞上清液的培养基测定最大LDH活性。实验是按照制造商的说明进行的。将一部分培养基用等体积的LDH底物溶液处理30分钟,然后用5体积的0.1M NaOH停止;使用分光光度计测量用1/100体积的10%Triton X-100处理并在37°C下孵育30分钟的姐妹培养物在440 nm处的吸光度。 Caspase-3和Caspase-9活性测定[1] 我们测量了产色胱天蛋白酶底物Ac-DEVD-pNA(乙酰基Asp-Glu-Val-Asp对硝基苯胺)、胱天蛋白酶-3底物Ac-LEHD-pNA和胱天蛋白酶-9底物Ac-LEHD-His-Asp对硝基苯胺的切割,分别计算胱天蛋白酶3和胱天酶-9的活性。实验按照制造商的协议进行。将约50µg的总蛋白加入含有Ac-DEVD-pNA(2 mM)或Ac-LEHD-pNA的反应缓冲液中,然后在37°C下孵育2小时。使用光谱仪在405nm下测量从其相应前体裂解的黄色pNA的吸光度。然后,将标准化为细胞裂解物总蛋白的特定胱天蛋白酶活性表示为相对于在含有10%FBS的DMEM中培养的对照细胞基线的倍数变化。 蛋白质印迹分析[1] 我们使用蛋白质裂解缓冲液以及适当浓度的蛋白酶抑制剂和磷酸酶抑制剂来提取细胞蛋白质。然后,使用Bradford测定法检测总蛋白浓度。蛋白质样品通过SDS-PAGE(12%聚丙烯酰胺凝胶)分级,然后转移到硝化纤维膜上。在含有5%脱脂奶粉的阻断缓冲液(20 mM Tris,pH 7.6,150 mM NaCl和0.1%吐温-20)中孵育后,将膜与指定抗体在4°C下孵育过夜,然后在室温下与辣根过氧化物酶(HRP)偶联的二抗反应1小时。使用增强的化学发光试剂通过化学发光检测试剂盒显示蛋白质条带。β-肌动蛋白被用作内部对照。 溶液制备。[3] 通过在硅化微量离心管中蒸发10mm乙醇溶液(在氮气流下)来制备HETE溶液,如12-HETE和其他亲脂性化合物。将二甲基亚砜(<最终体积的0.05%)加入乙醇中,以防止亲脂性物质完全干燥到管壁上。蒸发后,加入1ml培养基,使用高功率声波探针分散药物。特别注意确保溶液不会产生泡沫或摸起来很热。分散后,通过与适量的培养基混合,将所有溶液放入硅化玻璃管中,使其达到最终体积。 神经元培养。[3] 根据先前发表的方法制备原代皮质神经元培养物(Grimaldi和Cavallaro,1999)。简而言之,从怀孕17天的Wistar大鼠身上提取胎儿。然后解剖皮质,切成小块,在37°C下用木瓜蛋白酶(1mg/ml)孵育9分钟。在此之后,通过火抛光巴斯德移液管分离组织,并通过在Earle平衡盐溶液中以10mg/ml牛血清白蛋白(BSA)和10mg/ml卵粘液(胰蛋白酶抑制剂)组成的梯度离心分离所得细胞悬浮液。然后将沉淀物重新悬浮在高糖、无酚红的DMEM中,DMEM含有10%胎牛血清、2mm谷氨酰胺、100IU青霉素和100μg/ml链霉素。使用台盼蓝排斥试验对细胞进行计数和存活率测试;接下来,将3.2×105个细胞/孔接种到聚-d-赖氨酸涂层48孔板的内24孔上。板的外孔充满水,以减少培养物的蒸发。接种96小时后,加入含有10μm氟脱氧尿苷和10μm尿苷的混合物以阻断神经胶质细胞的生长。7天后,再次加入100μl含有神经胶质细胞抑制混合物的新鲜培养基,以补偿营养耗竭。 NMDA-R介导的毒性程序。[3] 皮质神经元在体外培养13-15天。在实验之前,将维持细胞的一半培养基移除并保留以备后用。通过将培养物暴露于200μm谷氨酸盐中15分钟,在由(以毫米计):125 NaCl、25葡萄糖、10 HEPES、pH 7.4、5 KCl和1.8氯化钙以及2.5%无脂肪酸BSA组成的无镁盐水溶液中检测谷氨酸盐毒性。暴露后,用生理盐水洗涤细胞两次,并在由70%原始培养基(神经元已在其中培养了13-15天)和30%新鲜培养基组成的培养基中孵育。然后将细胞孵育18小时,之后检查培养基中的乳酸脱氢酶(LDH)水平,并将其用作细胞毒性的指标。初步研究表明,500 nm MK-801可以预防谷氨酸毒性,证实了NMDA-R介导的机制(数据未显示)。 AMPA和红藻氨酸受体介导的毒性程序。[3] 为了检测AMPA和红藻氨酸受体介导的毒性,神经元培养6-9天,然后在分析前暴露于100μm谷氨酸盐和25μm环噻嗪(用于防止AMPA-R脱敏)和500 nm MK-801 18-20小时。 上述神经元制备技术导致主要的神经元培养,尽管仍有有限数量的星形胶质细胞存在。这种细胞类型对谷氨酸毒性具有抗性(Amin和Pearce,1997),尽管有报道称在环噻嗪存在的情况下会发生这种细胞死亡(David等人,1996)。初步研究证实,星形胶质细胞在我们的培养物中对AMPA-红藻氨酸毒性没有显著影响。当星形胶质细胞在与神经元制备相同的条件下暴露于谷氨酸并放置20小时时,LDH水平仅增加到背景的105%,而在富含神经元的培养物中为150-200%(数据未显示)。因此,可以得出结论,星形胶质细胞污染对我们神经元培养物中谷氨酸的影响没有实质性影响。 毒性试验。[3] 根据Decker和Lohmann-Matthes(1988)的方法,在损伤后18-20小时通过测量LDH释放到无酚红培养基中来评估细胞毒性。在每个点用四倍值进行实验,所有平板均含有谷氨酸盐(阳性)和非谷氨酸盐(阴性)对照。所需的测定开发时间因培养批次和体外天数而异,当阳性对照孔在495-650 nm处的吸光度值为0.3-0.4 au时记录值。该测定通过与线粒体功能存活率测定[2,3-双(2-甲氧基-4-硝基-5-磺苯基)-(2H)-四唑-5-甲酰苯胺(XTT)测定;Roehm等人,1991]进行比较进行了验证。两种系统获得的结果相似,尽管本研究中使用了LDH释放,因为它提供了比XTT测定更大的信噪比。阴性对照组和阳性对照组的平均LDH水平之比被用作培养质量的指标。不包括阴性对照中平均LDH水平超过最大值(阳性对照)60%的试验数据。 |
| 药代性质 (ADME/PK) |
Metabolism / Metabolites
12-HETE is a known human metabolite of arachidonic acid. |
| 毒性/毒理 (Toxicokinetics/TK) |
Pertussis toxin studies [3]
Pertussis toxin (PTx, an agent that inactivates Gi/o proteins) was used to investigate whether 12-HETE exerts its effect by direct action on N- and L-type VSCCs or by activating a Gi/o-protein-coupled receptor (Zamponi and Snutch, 1998; Kaneko et al., 1999). After treatment with 100 ng/ml PTx overnight, the anti-excitotoxic effect of 12-(S)HETE was eliminated (Fig.7), which suggests that 12-(S)HETE inhibits N- and L-type VSCCs via Gi/o-protein linkage rather than through a direct action. The effect of PTx on 12-(S)HETE-inhibited calcium influx was also examined by calcium imaging. As with the glutamate toxicity studies, PTx treatment eliminated the inhibitory effect of 12-HETE on glutamate-induced [Ca2+]i (Fig.8A,C), which suggests that 12-(S)HETE inhibits glutamate-stimulated calcium influx via a Gi/o-protein-coupled mechanism. |
| 参考文献 |
|
| 其他信息 |
(5Z,8Z,10E,14Z)-12-hydroxyicosatetraenoic acid is the (5Z,8Z,10E,14Z)-stereoisomer of 12-HETE. It has a role as a mouse metabolite. It is a conjugate acid of a 12-HETE(1-).
12-Hydroxy-5,8,10,14-eicosatetraenoic Acid is a metabolite generated from arachidonic acid by 12S-type arachidonate 12-lipoxygenase in platelets. 12-hydroxy-5,8,10,14-eicosatetraenoic acid (12(S)-HETE) may be involved in inflammation and pruritus. A lipoxygenase metabolite of ARACHIDONIC ACID. It is a highly selective ligand used to label mu-opioid receptors in both membranes and tissue sections. The 12-S-HETE analog has been reported to augment tumor cell metastatic potential through activation of protein kinase C. (J Pharmacol Exp Ther 1995; 274(3):1545-51; J Natl Cancer Inst 1994; 86(15):1145-51) Background: The dysfunction of cell apoptosis is an important event in the progression of cancer, and the growth of cancer cells is negatively regulated by cell apoptosis. In different types of cancers, inhibition of cellular apoptosis is often observed in the cancerous tissue, and increased resistance to apoptosis is a hallmark of cancer. Although previous studies have shown that 12-lipoxygenase (12-LOX)/12-hydroxyeicosatetraenoic acid (12-HETE) is activated and upregulated in different types of cancers, the consequences of 12-LOX/12-HETE upregulation and its precise roles in the survival of ovarian carcinoma cells are still unknown. Methods: MTT assays, caspase activity assays, lactate dehydrogenase (LDH) assays, and Western blot analysis were the methods used in this study. Our study suggests that 12-HETE inhibits the mitochondria-dependent apoptosis pathway through the ILK pathway and that NF-κB acts as a downstream target of 12-HETE-activated ILK. This study implies that 12-LOX/12-HETE and its downstream targets (ILK and NF-κB) play critical roles in promoting the survival of ovarian cancer cells and positively regulate the progression of ovarian cancer, which indicates a new potential target for future treatment of ovarian cancer. [1] This review reports the up to now knowledge about 12-HETE, in particular focusing on the role of the biosynthesis of this 12-LOX metabolite in platelets. The analysis of the literature reveals the presence of different analytical methods for the detection of eicosanoids, with good specificity, but the sample preparation still needs to be implemented. The majority of analytical techniques present in the literature are independent from the source of this AA metabolite, even if different cell types may differ in the kinetic of 12-HETE production. For this reason, who intends to actually study platelet 12-HETE from a physiological and clinical point of view cannot avoid taking into consideration the matrix in which the measurements should be performed. We think that serum might be the biological sample ideally suitable to this goal for several reasons. The handling of serum is simple, convenient and time saving because no steps are required for cell isolation. In the serum prostanoids produced during clotting are the result of the interaction between various cell types, simulating in vivo conditions. Even if this type of sample do not reflect basal concentrations, it constitutes a suitable and easy-to-obtain ex vivo system to investigate simultaneously the activity of both platelet COX and LOX enzymes. However, we believe that essential premise is to study in depth the kinetic of 12-HETE formation in platelets because some authors reported a continuous production of this molecule. The amount of 12-HETE produced from rat platelets aggregated by collagen increased continuously, even after 115 min as platelet 12-LOX is able to utilize, not only AA endogenously present in the cell, but also that one derived from plasma phospholipids. Even more important, this enzyme is resistant to inactivation. These data has been confirmed also by Sautebin in humans. So all these points must be taken into account in order to develop an analytical method for measurement of 12-HETE derived from platelets. As documented, the information on the physiological and pathological functions of this 12-LOX metabolite are so far not completely clarified. Conflicting results have been reported about the role of 12-HETE in platelet function. Indeed, whatever the agonist used to induce platelet aggregation, 12-HETE showed to exert both platelet pro- and anti-aggregant activity, with the exception of platelet aggregation induced by thrombin, in which exogenously added 12-HETE was shown to have proaggregatory activity. The increased platelet proaggregatory activity of 12-HETE may be of relevance in controlling the occurrence of thrombotic phenomena in human. In this context the observation that 12-HETE synthesis is involved in transient heparin-induced platelet activation during carotid endarterectomy is of particular relevance. So the understanding of mechanisms by which 12-LOX and its metabolites modulate platelet function might be of help not only in elucidating the role of this 12-hydroxy fatty acid in the thrombotic process, but also may provide the rationale for the development of a novel class of anti-platelet drugs. [2] While the importance of cyclooxygenase (COX) in platelet function has been amply elucidated, the identification of the role of 12-lipoxygenase (12-LOX) and of its stable metabolite, 12-hydroxyeicosatretraenoic acid (12-HETE), has not been clarified as yet. Many studies have analysed the implications of 12-LOX products in different pathological disorders but the information obtained from these works is controversial. Several analytical methods have been developed over the years to simultaneously detect eicosanoids, and specifically 12-HETE, in different biological matrices, essentially enzyme-linked immunosorbent assays (ELISA), radioimmunoassays (RIA), high performance liquid chromatography (HPLC) and mass spectrometry coupled with both gas and liquid chromatography methods (GC- and LC-MS). This review is aimed at summarizing the up to now known physiological and clinical features of 12-HETE together with the analytical methods used for its determination, focusing on the critical issues regarding its measurement. [2] 12-hydroxyeicosatetraenoic acid (12-HETE) is a neuromodulator that is synthesized during ischemia. Its neuronal effects include attenuation of calcium influx and glutamate release as well as inhibition of AMPA receptor (AMPA-R) activation. Because 12-HETE reduces ischemic injury in the heart, we examined whether it can also reduce neuronal excitotoxicity. When treated with 12-(S)HETE, cortical neuron cultures subjected to AMPA-R-mediated glutamate toxicity suffered up to 40% less damage than untreated cultures. The protective effect of 12-(S)HETE was concentration-dependent (EC50 = 88 nm) and stereostructurally selective. Maximal protection was conferred by 300 nm 12-(S)HETE; 300 nm 15-(S)HETE was similarly protective, but 300 nm 5-(S)HETE was less effective. The chiral isomer 12-(R)HETE offered no protection; neither did arachidonic acid or 12-(S)hydroperoxyeicosatetraenoic acid. Excitotoxicity was calcium-dependent, and 12-(S)HETE was demonstrated to protect by inactivating N and L (but not P) calcium channels via a pertussis toxin-sensitive mechanism. Calcium imaging demonstrated that 12-(S)HETE also attenuates glutamate-induced calcium influx into neurons via a pertussis toxin-sensitive mechanism, suggesting that it acts via a G-protein-coupled receptor. In addition, 12-(S)HETE stimulates GTPgammaS binding (indicating G-protein activation) and inhibits adenylate cyclase in forskolin-stimulated cultures over the same concentration range as it exerts its anti-excitotoxic and calcium-influx attenuating effects. These studies demonstrate that 12-(S)HETE can protect neurons from excitotoxicity by activating a G(i/o)-protein-coupled receptor, which limits calcium influx through voltage-gated channels.[3] During the drafting of this manuscript a report was published indicating that 12-(S)HETE has some affinity for the low-affinity leukotriene B4 receptor (BLT2) (Yokomizo et al., 2001). However, the features of BLT2 are considerably different from those described here; in particular, this receptor is not pertussis-toxin sensitive, 12-(S)HETE binds to BLT2 with an EC50 in the micromolar rather than nanomolar range, and most importantly, mRNA to BLT2 is found in almost every tissue except the brain (Yokomizo et al., 2000). In summary, the data presented in this study demonstrate that 12-(S)HETE significantly attenuates neuronal AMPA toxicity by reducing calcium influx through N- and L-type calcium channels during glutamate exposure. The protective effect of 12-HETE is stereostructurally defined and is mediated via a PTx-sensitive Gi/o protein linkage. As with other Gi/o-protein-coupled receptor systems, the putative 12-(S)HETE receptor inhibits adenylate cyclase activity in addition to inhibiting VSCCs. The concentration range over which 12-(S)HETE inhibits cAMP formation and stimulates GTPγS binding is similar to that over which it reduces glutamate toxicity. Together, these data strongly support the existence of a G-protein-coupled 12-(S)HETE receptor in rat cortical neuronal membranes, which may act as a neuroprotective system during excitotoxic events such as ischemia.[3] |
| 分子式 |
C20H32O3
|
|---|---|
| 分子量 |
320.46628
|
| 精确质量 |
320.235
|
| 元素分析 |
C, 74.96; H, 10.07; O, 14.98
|
| CAS号 |
71030-37-0
|
| 相关CAS号 |
12-HETE-d8;2525175-25-9
|
| PubChem CID |
13786989
|
| 外观&性状 |
Colorless to light yellow liquid
|
| 密度 |
1.0±0.1 g/cm3
|
| 沸点 |
487.7±45.0 °C at 760 mmHg
|
| 闪点 |
262.8±25.2 °C
|
| 蒸汽压 |
0.0±2.8 mmHg at 25°C
|
| 折射率 |
1.514
|
| LogP |
5.45
|
| tPSA |
17.07
|
| 氢键供体(HBD)数目 |
2
|
| 氢键受体(HBA)数目 |
3
|
| 可旋转键数目(RBC) |
14
|
| 重原子数目 |
23
|
| 分子复杂度/Complexity |
392
|
| 定义原子立体中心数目 |
0
|
| SMILES |
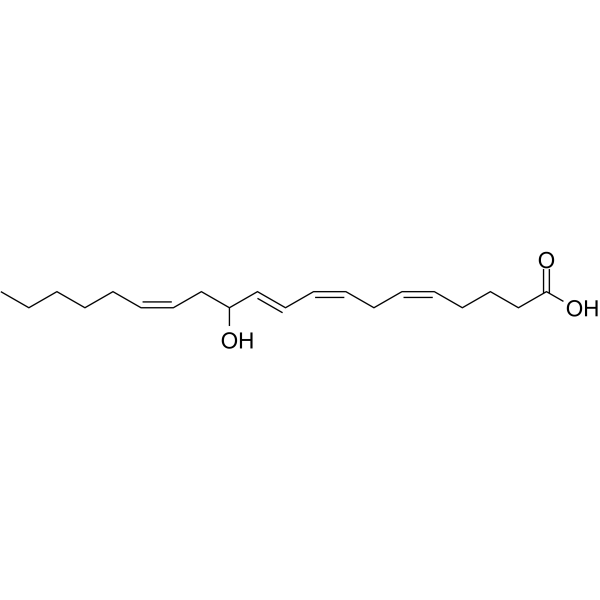
CCCCC/C=C\CC(/C=C/C=C\C/C=C\CCCC(=O)O)O
|
| InChi Key |
ZNHVWPKMFKADKW-VXBMJZGYSA-N
|
| InChi Code |
InChI=1S/C20H32O3/c1-2-3-4-5-10-13-16-19(21)17-14-11-8-6-7-9-12-15-18-20(22)23/h7-11,13-14,17,19,21H,2-6,12,15-16,18H2,1H3,(H,22,23)/b9-7-,11-8-,13-10-,17-14+
|
| 化学名 |
(5Z,8Z,10E,14Z)-12-hydroxyicosa-5,8,10,14-tetraenoic acid
|
| 别名 |
71030-37-0; 12-Hete; 5,8,10,14-Eicosatetraenoicacid, 12-hydroxy-, (5Z,8Z,10E,14Z)-; 12-hydroxy-5Z,8Z,10E,14Z-eicosatetraenoic acid; (5Z,8Z,10E,14Z)-12-hydroxyicosatetraenoic acid; (+/-)12-HETE; 12-Hydroxyeicosatetraenoic acid; (5Z,8Z,10E,14Z)-12-hydroxyicosa-5,8,10,14-tetraenoic acid;
|
| HS Tariff Code |
2934.99.9001
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
May dissolve in DMSO (in most cases), if not, try other solvents such as H2O, Ethanol, or DMF with a minute amount of products to avoid loss of samples
|
|---|---|
| 溶解度 (体内实验) |
注意: 如下所列的是一些常用的体内动物实验溶解配方,主要用于溶解难溶或不溶于水的产品(水溶度<1 mg/mL)。 建议您先取少量样品进行尝试,如该配方可行,再根据实验需求增加样品量。
注射用配方
注射用配方1: DMSO : Tween 80: Saline = 10 : 5 : 85 (如: 100 μL DMSO → 50 μL Tween 80 → 850 μL Saline)(IP/IV/IM/SC等) *生理盐水/Saline的制备:将0.9g氯化钠/NaCl溶解在100 mL ddH ₂ O中,得到澄清溶液。 注射用配方 2: DMSO : PEG300 :Tween 80 : Saline = 10 : 40 : 5 : 45 (如: 100 μL DMSO → 400 μL PEG300 → 50 μL Tween 80 → 450 μL Saline) 注射用配方 3: DMSO : Corn oil = 10 : 90 (如: 100 μL DMSO → 900 μL Corn oil) 示例: 以注射用配方 3 (DMSO : Corn oil = 10 : 90) 为例说明, 如果要配制 1 mL 2.5 mg/mL的工作液, 您可以取 100 μL 25 mg/mL 澄清的 DMSO 储备液,加到 900 μL Corn oil/玉米油中, 混合均匀。 View More
注射用配方 4: DMSO : 20% SBE-β-CD in Saline = 10 : 90 [如:100 μL DMSO → 900 μL (20% SBE-β-CD in Saline)] 口服配方
口服配方 1: 悬浮于0.5% CMC Na (羧甲基纤维素钠) 口服配方 2: 悬浮于0.5% Carboxymethyl cellulose (羧甲基纤维素) 示例: 以口服配方 1 (悬浮于 0.5% CMC Na)为例说明, 如果要配制 100 mL 2.5 mg/mL 的工作液, 您可以先取0.5g CMC Na并将其溶解于100mL ddH2O中,得到0.5%CMC-Na澄清溶液;然后将250 mg待测化合物加到100 mL前述 0.5%CMC Na溶液中,得到悬浮液。 View More
口服配方 3: 溶解于 PEG400 (聚乙二醇400) 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 3.1204 mL | 15.6021 mL | 31.2042 mL | |
| 5 mM | 0.6241 mL | 3.1204 mL | 6.2408 mL | |
| 10 mM | 0.3120 mL | 1.5602 mL | 3.1204 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
 Degarelix acetate hydrate
Degarelix acetate hydrate
 DDO-2728
DDO-2728
 Vatalanib hydrochloride (Vatalanib hydrochloride; PTK787 hydrochloride; ZK-222584 hydrochloride; CGP-797870 hydrochloride)
Vatalanib hydrochloride (Vatalanib hydrochloride; PTK787 hydrochloride; ZK-222584 hydrochloride; CGP-797870 hydrochloride)
 Tuspetinib hydrate (HM43239 hydrate)
Tuspetinib hydrate (HM43239 hydrate)
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1
































 463611831
463611831
