| 规格 | 价格 | 库存 | 数量 |
|---|---|---|---|
| 1mg |
|
||
| 5mg |
|
||
| 10mg |
|
||
| 25mg |
|
||
| 50mg |
|
||
| 100mg |
|
||
| 250mg |
|
||
| Other Sizes |
|
| 靶点 |
Rac GTPase (IC50 = 50 μM)
NSC 23766 mainly targets Rac1 GTPase, inhibiting Tiam1 (a guanine nucleotide exchange factor for Rac1)-Rac1 interaction to block Rac1 activation; the IC50 value for human recombinant Rac1 is 100 nM [1] NSC 23766 has no obvious inhibitory effect on other Rho family GTPases (Cdc42, RhoA) (IC50>1000 nM), showing high target specificity [2] |
|---|---|
| 体外研究 (In Vitro) |
NSC23766 经鉴定适合 Rac1 的表面凹槽,已知该凹槽对 GEF 规范至关重要。 NSC23766 以剂量依赖性方式有效抑制 Rac 特异性 GEF Trio 或 Tiam1 的结合和激活,而不干扰密切相关的 Cdc42 或 RhoA 各自 GEF 的结合或激活,也不干扰 Rac1 与 BcrGAP 或效应器 PAK1 的相互作用。 NSC 23766 积极调节细胞骨架上的 Rac GTPase 功能和许多细胞功能,包括细胞周期、细胞生长、粘附、迁移和基因转录。 NSC 23766 (50 μM) 有效阻断血清或血小板衍生生长因子诱导的 Rac1 激活和板状伪足形成,而不影响 NIH 3T3 细胞中内源性 Cdc42 或 RhoA 的活性。 NSC 23766 减少 Trio 或 Tiam1,但不减少 Vav、Lbc、Intersectin 或组成型活性 Rac1 突变体刺激的 NIH 3T3 细胞生长,并抑制 Trio、Tiam1 或 Ras 诱导的细胞转化。 NSC23766 剂量依赖性地抑制 PC-3 细胞增殖和贴壁依赖性生长。 25 μM NSC23766 可抑制 PC-3 细胞通过 Matrigel 的侵袭 85%。 [1] 50 μM NSC 23766 抑制凝血酶诱导的人血小板中 Rac1 和 Rac2 的激活以及血小板聚集。 NSC23766 可阻止 swAPP-HEK293 细胞中 Aβ40 和 Aβ42 的产生,而不影响 Notch 和 sAPPα。 NSC23766 可阻止细胞中的 γ-分泌酶活性,但不能作为直接的 γ-分泌酶抑制剂。 NSC23766 剂量依赖性地降低分泌型和细胞内 Aβ40 的水平,IC50 为 48.94 μM。 50 μM NSC 23766 可抑制 Aβ42 的释放 57.97%。 NSC23766 调节内皮一氧化氮合酶表达和内皮功能。 100 μM NSC23766 在牛主动脉 EC 中抑制 eNOS 启动子活性 60%,在 bEND.3 细胞中抑制 30% 至 35%。 NSC23766 抑制 Rac1 会破坏 eNOS mRNA 的稳定性,并将其半衰期缩短至 17 小时。 NSC23766 剂量依赖性地减弱 ACh 诱导的野生型小鼠主动脉环松弛。 NSC23766 抑制细胞生长并诱导细胞凋亡。 NSC23766 以剂量依赖性方式降低 MDA-MB-468 和 MDA-MB-231 细胞活力,IC50 约为 10 μM,与雌激素受体 (ER)、孕激素受体 (PR)、Her2、和p53突变。 NSC23766对MCF12A正常乳腺上皮细胞的存活影响很小。暴露于 NSC 23766 24 小时后,MDA-MB-231 细胞的 G1 期细胞数量从 41% 增加至 65%,同时 S 期和 G2-M 期细胞数量减少。 100 μM NSC23766 诱导凋亡 MDA-MB-468 增加六倍。 NSC23766 对乳腺癌细胞细胞周期停滞或凋亡的抑制作用是通过下调细胞周期蛋白 D1、生存素和 X 连锁蛋白凋亡抑制剂介导的。激酶测定:细胞在 10 cm 培养皿中以对数期生长,并在 0.5% 血清培养基中饥饿或以其他方式指示 24 小时,然后在含有 20 mM Tris HCl (pH 7.6)、100 mM NaCl、10 的缓冲液中裂解。 mM MgCl2、1% Nonidet P-40、10% 甘油和 1× 蛋白酶抑制剂混合物。澄清裂解物,标准化蛋白质浓度,并通过效应域下拉测定法测量裂解物中 GTP 结合的 Rac1。对于 His6-PAK1 PBD Pull-down 测定,细胞裂解物与从大肠杆菌中纯化的 Ni2+-琼脂糖固定的 His6-PAK1 PBD 结构域(每个约 1 μg)一起孵育 30 分钟。 Ni2+-琼脂糖共沉淀物在洗涤缓冲液中洗涤两次,并使用抗 Rac1 单克隆抗体进行免疫印迹分析。细胞测定:将细胞(1.5 × 104/mL 人乳腺癌细胞 MDA-MB-468)接种到含有 200 μL 培养基的 96 孔组织培养板的每个孔中。铺板 24 小时后,用 200 μL 含有所示浓度 NSC23766 的新鲜培养基更换培养基。处理结束时,每孔加入20μL MTS溶液,37℃孵育2小时。在 96 孔板读数器上读取 490 nm 处的吸光度。
NSC 23766 浓度依赖性抑制Rac1 GTP酶活性:10 μM浓度下,小鼠巨噬细胞中Rac1活性降低65%;50 μM浓度下,猪卵母细胞中Rac1活性降低78%,且不影响Cdc42、RhoA活性 [2][4] NSC 23766 抑制猪卵母细胞成熟:100 μM浓度处理44小时,第一极体排出率从72%降至28%,同时下调Cyclin B1表达(降低55%)、上调p21表达(升高2.3倍)[2] NSC 23766 影响小鼠原始卵泡形成:50 μM浓度处理卵巢颗粒细胞24小时,STAT3磷酸化水平降低60%,Jagged1、GDF9、BMP15的mRNA表达分别降低58%、52%、49% [3] NSC 23766 抑制炎症相关因子分泌:50 μM浓度处理LPS刺激的巨噬细胞,TNF-α、IL-6分泌量分别降低62%、57%,NF-κB p65核转位减少55% [4] NSC 23766 调节免疫细胞功能:50 μM浓度处理小鼠脾淋巴细胞,CD4+ T细胞增殖率从68%降至32%,IFN-γ分泌减少65%,IL-4分泌升高2.1倍 [1] |
| 体内研究 (In Vivo) |
NSC23766 诱导造血干细胞/祖细胞的动员。将 NSC23766 (2.5 mg/kg) 腹膜内施用到“活动性差”的 C57Bl/6 小鼠品系中,导致注射后 6 小时循环造血干细胞祖细胞增加两倍。 NSC23766 可减轻脂多糖诱导的小鼠急性肺损伤。 NSC23766以1或3mg/kg治疗不仅减少炎症细胞浸润和MPO活性,而且抑制促炎介质、肿瘤坏死因子-α和白介素-1β的mRNA表达。 NSC23766 还可以减少 LPS 挑战的肺部中伊文思蓝和白蛋白的积累。
NSC 23766 以50 mg/kg剂量腹腔注射,每周3次,持续12周,可显著预防NOD小鼠1型糖尿病发生:发病率从85%降至30%,胰腺胰岛炎评分从3.2分降至1.1分,胰岛CD4+ T细胞浸润减少70% [1] NSC 23766 腹腔注射30 mg/kg,可缓解蜜蜂毒液诱导的小鼠急性炎症痛:给药后2小时,机械痛阈从0.8 g升至2.5 g,热痛阈从12.5秒升至21.3秒,脊髓组织中Rac1活性降低68% [4] NSC 23766 以20 mg/kg腹腔注射,每日1次,持续7天,可减少新生小鼠卵巢原始卵泡数量:从对照组的126个/卵巢降至78个/卵巢,STAT3磷酸化水平降低52% [3] |
| 酶活实验 |
使用蛋白酶和磷酸酶抑制剂匀浆来自腰椎增大的新鲜脊髓组织,然后使用缓冲液裂解组织。收集上清液,并在 4°C 下以 12,000×g 离心 5 分钟后,与 PAK-PBD 珠子在 4°C 下在旋转器上孵育 1 小时。然后以 5000×g 离心 3 分钟,将珠子沉淀。在 4°C 下。在 LaemmLi 缓冲液中重悬后,将所得沉淀煮沸两分钟。对珠子样品进行蛋白质印迹分析。蛋白质印迹分析还用于确定每个样品中的总 Rac1。
采用GST-pull-down法检测Rac1活性:提取细胞或组织总蛋白,与GST-PAK1-PBD融合蛋白(特异性结合活化态Rac1-GTP)孵育,4℃孵育1小时后,洗涤沉淀并进行Western blot检测,定量分析活化态Rac1的相对含量,计算NSC 23766对Rac1活性的抑制率 [2] 采用荧光共振能量转移(FRET)法验证结合特异性:将重组Rac1与荧光标记GTP混合,加入梯度浓度的NSC 23766,37℃孵育30分钟,检测FRET信号变化,计算药物与Rac1的结合亲和力及IC50值 [4] |
| 细胞实验 |
96 孔组织培养板每孔接种 1.5 × 10 4 /mL 细胞和 200 μL 培养基。 24 小时电镀期后,用 200 μL 含有指定浓度 NSC23766 的新培养基替换培养基。处理期结束后,将 20 μL MTS 溶液引入每个孔中,并在 37°C 下孵育两小时。使用 96 孔板读数器测量 490 nm 处的吸光度。
猪卵母细胞分离自卵巢,接种于含M199培养基的培养皿,38.5℃、5%CO2培养2小时后,加入梯度浓度的NSC 23766(10、50、100 μM),继续培养44小时;观察第一极体排出情况,提取蛋白通过Western blot检测Rac1、Cdc42、RhoA活性及Cyclin B1、p21表达 [2] 小鼠卵巢颗粒细胞分离后接种于24孔板(1×10⁵个/孔),DMEM/F12培养基培养24小时,加入NSC 23766(20、50、100 μM)孵育24小时;提取总RNA,qPCR检测Jagged1、GDF9、BMP15的mRNA表达;提取蛋白检测STAT3磷酸化水平 [3] 小鼠巨噬细胞接种于96孔板(5×10⁴个/孔),培养16小时后,加入LPS(1 μg/mL)刺激,同步加入NSC 23766(10、50 μM),孵育24小时;ELISA法检测细胞上清中TNF-α、IL-6浓度,GST-pull-down法检测Rac1活性 [4] |
| 动物实验 |
7周龄时,将Balb/c对照组和NOD小鼠随机分为四组(每组n=8)。8周龄时,两组实验组(Balb/c小鼠和NOD小鼠)接受NSC23766(2.5 mg/kg/天,腹腔注射),而另外两组(Balb/c对照组和NOD小鼠)接受等体积的生理盐水注射。在接下来的34周内,每周测量血糖和体重。两组实验动物(Balb/c小鼠和NOD小鼠)接受NSC23766治疗,而两组对照组接受等体积的生理盐水注射。连续34周每周测量体重和血糖。采用GLISA激活试验检测胰岛中Rac1的激活情况。采用蛋白质印迹法检测Rac1和CHOP的表达。[1]
将NOD小鼠(雌性,4周龄)随机分为两组:给药组腹腔注射溶于5% DMSO+95%生理盐水的NSC 23766(50 mg/kg),每周三次,持续12周;对照组注射等体积的溶剂。每2周检测一次血糖(≥11.1 mmol/L诊断为糖尿病),并在实验结束时收集胰腺组织进行HE染色和免疫组化,以分析胰岛炎的严重程度和T细胞浸润。[1] 采用ICR小鼠(雄性,6周龄)建立蜂毒诱导的急性炎症疼痛模型:将蜂毒(20 μL/只)皮下注射到右后爪,30分钟后给药;将NSC 23766溶于生理盐水中,并以10和30 mg/kg的剂量进行腹腔注射。分别于给药后1、2和4小时检测机械痛阈(von Frey纤维法)和热痛阈(热板法),并在实验结束时检测脊髓和背根神经节中的Rac1活性[4]。新生C57BL/6小鼠(出生后1天)随机分为两组:给药组腹腔注射NSC 23766,剂量为20 mg/kg,每日一次,连续7天;对照组注射生理盐水。实验结束时收集卵巢组织,通过HE染色计数原始卵泡数量,并通过Western blot检测STAT3磷酸化水平[3]。 |
| 毒性/毒理 (Toxicokinetics/TK) |
NSC 23766在体外浓度≤100 μM时未显示明显的细胞毒性,细胞存活率≥80%[2][3]
当以50 mg/kg的剂量腹腔注射NSC 23766(每周三次,持续12周)时,NOD小鼠体重增长正常,血清ALT、AST、BUN、Cr水平与对照组相比无统计学差异,且胰腺、肝脏或肾脏未见明显的病理损伤[1] 小鼠单次腹腔注射NSC 23766的最大耐受剂量≥100 mg/kg,未观察到急性毒性[4] |
| 参考文献 |
|
| 其他信息 |
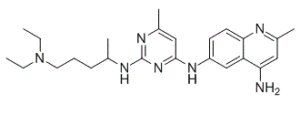
NSC 23766 是一种氨基嘧啶类化合物,其结构为 6-甲基嘧啶-2,4-二胺,其中 2 位和 4 位上的氨基分别被 5-(二乙氨基)戊-2-基和 4-氨基-2-甲基喹啉-6-基取代。它是一种信号 G 蛋白 RAC1(Ras 相关 C3 肉毒杆菌毒素底物 1)的抑制剂。NSC 23766 具有多种活性,包括 EC 3.6.5.2(小单体 GTP 酶)抑制剂、抗病毒剂、毒蕈碱受体拮抗剂和细胞凋亡诱导剂。它是一种氨基喹啉、氨基嘧啶、伯氨基化合物、仲氨基化合物和叔氨基化合物。
背景/目的:1 型糖尿病 (T1D) 的特征是由于入侵的免疫细胞释放的细胞因子(例如白细胞介素-1β;IL-1β)破坏胰岛 β 细胞而导致的绝对胰岛素缺乏。这些细胞因子诱导 β 细胞功能障碍的机制仍不甚明了。最近的证据表明,吞噬细胞样 NADPH 氧化酶 2 (Nox2) 过度产生活性氧 (ROS),以及 β 细胞中抗氧化剂水平显著降低,会促使 β 细胞发生氧化损伤。Rac1 是一种小 G 蛋白,是 Nox2 全酶的成员之一。我们近期报道,Rac1 的已知抑制剂 NSC23766 可显著减弱体外胰岛β细胞中细胞因子诱导的 Nox2 活化和 ROS 生成。本研究旨在探讨 NSC23766(2.5 mg/kg/天,腹腔注射)对 NOD 小鼠(一种 1 型糖尿病模型)糖尿病发生发展的影响。方法:将实验动物分为两组(Balb/c 小鼠和 NOD 小鼠),分别给予 NSC23766 和等体积的生理盐水作为对照。每周测量小鼠体重和血糖,持续 34 周。采用 GLISA 活化检测法检测胰岛中 Rac1 的活化情况。采用蛋白质印迹法检测 Rac1 和 CHOP 的表达。结果:我们的研究结果表明,给予 NSC23766 可显著预防 NOD 小鼠自发性糖尿病的发生。此外,NSC23766显著抑制了NOD小鼠胰岛中Rac1的表达和活性以及内质网应激(CHOP表达)。结论:我们的研究结果首次证实了Tiam1-Rac1-Nox2信号通路在NOD小鼠自发性糖尿病发生中的作用。[1]哺乳动物卵母细胞的不对称分裂依赖于纺锤体的偏心定位,从而形成极体。小信号G蛋白Rac1是GTP酶家族的成员,它调控多种细胞事件,包括细胞生长控制、细胞骨架重组和蛋白激酶的激活。然而,Rac1对猪卵母细胞成熟和早期胚胎发育的影响尚未完全阐明。本研究探讨了Rac1在卵母细胞成熟和胚胎卵裂中的作用。我们首先发现Rac1定位于猪卵母细胞的皮层,用NSC 23766处理抑制Rac1活性会导致极体排出失败。此外,大多数经处理的卵母细胞表现出纺锤体形态异常,表明Rac1可能参与猪卵母细胞纺锤体的形成。这可能是由于Rac1对MAPK的调控作用,因为NSC 23766处理后p-MAPK的表达降低。此外,我们发现经处理的卵母细胞中大多数减数分裂纺锤体的位置远离皮层,表明Rac1在减数分裂纺锤体的定位中发挥作用。我们的结果还表明,抑制Rac1活性会导致早期胚胎发育失败。因此,我们的研究揭示了Rac1 GTP酶在猪卵母细胞成熟和早期胚胎卵裂中的关键作用。[2] NSC 23766是首个Tiam1-Rac1信号通路特异性抑制剂,它通过阻断Tiam1-Rac1相互作用来抑制Rac1活化,从而调节免疫反应、炎症信号、细胞周期和生殖细胞发育。[1] 临床前研究表明,NSC 23766在治疗自身免疫性疾病(1型糖尿病)和急性炎症性疼痛方面具有潜在价值,并且可以调节卵巢原始卵泡的形成,可作为生殖相关研究的工具药物。[1][3][4] NSC 23766的作用机制涉及多个下游信号通路,包括STAT3、NF-κB和细胞周期调控通路,具有多效性但靶向性强。特异性[2][3][4] |
| 分子式 |
C24H35N7
|
|
|---|---|---|
| 分子量 |
421.58
|
|
| 精确质量 |
421.29539415
|
|
| CAS号 |
733767-34-5
|
|
| 相关CAS号 |
NSC 23766 trihydrochloride;1177865-17-6
|
|
| PubChem CID |
409805
|
|
| 外观&性状 |
Off-white to light yellow solid powder
|
|
| 密度 |
1.16g/cm3
|
|
| 沸点 |
632.4ºC at 760 mmHg
|
|
| 闪点 |
336.2ºC
|
|
| 折射率 |
1.646
|
|
| LogP |
8.023
|
|
| tPSA |
91.99
|
|
| 氢键供体(HBD)数目 |
3
|
|
| 氢键受体(HBA)数目 |
7
|
|
| 可旋转键数目(RBC) |
10
|
|
| 重原子数目 |
31
|
|
| 分子复杂度/Complexity |
514
|
|
| 定义原子立体中心数目 |
0
|
|
| SMILES |
NC1=CC(C)=NC2=CC=C(NC3=NC(NC(C)CCCN(CC)CC)=NC(C)=C3)C=C12
|
|
| InChi Key |
DEFBCZWQLILOJF-UHFFFAOYSA-N
|
|
| InChi Code |
InChI=1S/C24H35N7/c1-6-31(7-2)12-8-9-16(3)27-24-28-18(5)14-23(30-24)29-19-10-11-22-20(15-19)21(25)13-17(4)26-22/h10-11,13-16H,6-9,12H2,1-5H3,(H2,25,26)(H2,27,28,29,30)
|
|
| 化学名 |
6-N-[2-[5-(diethylamino)pentan-2-ylamino]-6-methylpyrimidin-4-yl]-2-methylquinoline-4,6-diamine
|
|
| 别名 |
|
|
| HS Tariff Code |
2934.99.9001
|
|
| 存储方式 |
Powder -20°C 3 years 4°C 2 years In solvent -80°C 6 months -20°C 1 month |
|
| 运输条件 |
Room temperature (This product is stable at ambient temperature for a few days during ordinary shipping and time spent in Customs)
|
| 溶解度 (体外实验) |
|
|||
|---|---|---|---|---|
| 溶解度 (体内实验) |
注意: 如下所列的是一些常用的体内动物实验溶解配方,主要用于溶解难溶或不溶于水的产品(水溶度<1 mg/mL)。 建议您先取少量样品进行尝试,如该配方可行,再根据实验需求增加样品量。
注射用配方
注射用配方1: DMSO : Tween 80: Saline = 10 : 5 : 85 (如: 100 μL DMSO → 50 μL Tween 80 → 850 μL Saline)(IP/IV/IM/SC等) *生理盐水/Saline的制备:将0.9g氯化钠/NaCl溶解在100 mL ddH ₂ O中,得到澄清溶液。 注射用配方 2: DMSO : PEG300 :Tween 80 : Saline = 10 : 40 : 5 : 45 (如: 100 μL DMSO → 400 μL PEG300 → 50 μL Tween 80 → 450 μL Saline) 注射用配方 3: DMSO : Corn oil = 10 : 90 (如: 100 μL DMSO → 900 μL Corn oil) 示例: 以注射用配方 3 (DMSO : Corn oil = 10 : 90) 为例说明, 如果要配制 1 mL 2.5 mg/mL的工作液, 您可以取 100 μL 25 mg/mL 澄清的 DMSO 储备液,加到 900 μL Corn oil/玉米油中, 混合均匀。 View More
注射用配方 4: DMSO : 20% SBE-β-CD in Saline = 10 : 90 [如:100 μL DMSO → 900 μL (20% SBE-β-CD in Saline)] 口服配方
口服配方 1: 悬浮于0.5% CMC Na (羧甲基纤维素钠) 口服配方 2: 悬浮于0.5% Carboxymethyl cellulose (羧甲基纤维素) 示例: 以口服配方 1 (悬浮于 0.5% CMC Na)为例说明, 如果要配制 100 mL 2.5 mg/mL 的工作液, 您可以先取0.5g CMC Na并将其溶解于100mL ddH2O中,得到0.5%CMC-Na澄清溶液;然后将250 mg待测化合物加到100 mL前述 0.5%CMC Na溶液中,得到悬浮液。 View More
口服配方 3: 溶解于 PEG400 (聚乙二醇400) 请根据您的实验动物和给药方式选择适当的溶解配方/方案: 1、请先配制澄清的储备液(如:用DMSO配置50 或 100 mg/mL母液(储备液)); 2、取适量母液,按从左到右的顺序依次添加助溶剂,澄清后再加入下一助溶剂。以 下列配方为例说明 (注意此配方只用于说明,并不一定代表此产品 的实际溶解配方): 10% DMSO → 40% PEG300 → 5% Tween-80 → 45% ddH2O (或 saline); 假设最终工作液的体积为 1 mL, 浓度为5 mg/mL: 取 100 μL 50 mg/mL 的澄清 DMSO 储备液加到 400 μL PEG300 中,混合均匀/澄清;向上述体系中加入50 μL Tween-80,混合均匀/澄清;然后继续加入450 μL ddH2O (或 saline)定容至 1 mL; 3、溶剂前显示的百分比是指该溶剂在最终溶液/工作液中的体积所占比例; 4、 如产品在配制过程中出现沉淀/析出,可通过加热(≤50℃)或超声的方式助溶; 5、为保证最佳实验结果,工作液请现配现用! 6、如不确定怎么将母液配置成体内动物实验的工作液,请查看说明书或联系我们; 7、 以上所有助溶剂都可在 Invivochem.cn网站购买。 |
| 制备储备液 | 1 mg | 5 mg | 10 mg | |
| 1 mM | 2.3720 mL | 11.8601 mL | 23.7203 mL | |
| 5 mM | 0.4744 mL | 2.3720 mL | 4.7441 mL | |
| 10 mM | 0.2372 mL | 1.1860 mL | 2.3720 mL |
1、根据实验需要选择合适的溶剂配制储备液 (母液):对于大多数产品,InvivoChem推荐用DMSO配置母液 (比如:5、10、20mM或者10、20、50 mg/mL浓度),个别水溶性高的产品可直接溶于水。产品在DMSO 、水或其他溶剂中的具体溶解度详见上”溶解度 (体外)”部分;
2、如果您找不到您想要的溶解度信息,或者很难将产品溶解在溶液中,请联系我们;
3、建议使用下列计算器进行相关计算(摩尔浓度计算器、稀释计算器、分子量计算器、重组计算器等);
4、母液配好之后,将其分装到常规用量,并储存在-20°C或-80°C,尽量减少反复冻融循环。
计算结果:
工作液浓度: mg/mL;
DMSO母液配制方法: mg 药物溶于 μL DMSO溶液(母液浓度 mg/mL)。如该浓度超过该批次药物DMSO溶解度,请首先与我们联系。
体内配方配制方法:取 μL DMSO母液,加入 μL PEG300,混匀澄清后加入μL Tween 80,混匀澄清后加入 μL ddH2O,混匀澄清。
(1) 请确保溶液澄清之后,再加入下一种溶剂 (助溶剂) 。可利用涡旋、超声或水浴加热等方法助溶;
(2) 一定要按顺序加入溶剂 (助溶剂) 。
|
|---|
|
|
 Fluorne rhodamine picolyl azide
Fluorne rhodamine picolyl azide
 Rhodamine picolyl azide
Rhodamine picolyl azide
 Rhodiolgin
Rhodiolgin
 K-Ras inhibitor 12
K-Ras inhibitor 12
InvivoChem的所有产品仅用于作科学研究,不面向患者销售
Copyright 2020 InvivoChem LLC | All Rights Reserved 粤ICP备20063088号-1
































 COA
COA



 463611831
463611831
